
Journal of Resources and Ecology >
Ecology and Diversity of Bracket Fungi in Oak Dominated Forests of Nepal, Central Himalaya
Received date: 2023-12-07
Accepted date: 2024-10-30
Online published: 2025-01-21
Bracket fungi (Polypores) are a morphologically distinct category of Agaricomycetes (Basidiomycota) that are important agents for decomposition and nutrient cycling in forest ecosystems. The species composition and ecology of these fungi in particular forest ecosystems are overlooked in Nepal. Thus, the current study is aimed at assessing the diversity and distribution of bracket fungi in oak-dominated forests in the middle mountains of central Nepal. Regular field visits over a six-year period (2017-2022) were done at nine study sites of oak-dominated forests in Karnabhumi Community Forest (KCF) and Gumalchoki Community Forest (GCF) from Makawanpur and Kathmandu districts respectively. A total of 26 species of bracket fungi belonging to 17 genera and 8 families were reported. The majority of the bracket fungi are known to be saprobic, while Coltricia cinnamomea is the only mushroom found to be mycorrhizal based on its ecology. Most of the bracket fungi were found to be inedible, while Ganoderma lucidum, Laetiporus sulphureus, and Pycnoporus cinnabarinus were found to be medicinal. The diversity indices, such as the Shannon Weiner index and the Simpson diversity index, were found to be 0.26 and 0.53 in GCF, respectively, and 0.17 and 0.35 in GCF. This suggest that Oak dominated forests harbor considerable diversity of bracket fungi such that conservation of these forests is necessary.
Key words: ectomycorrhizal fungi; hardwood fungi; polypores; Quercus forest; shelf fungi; wild mushroom
ADHIKARI Hari Sharan . Ecology and Diversity of Bracket Fungi in Oak Dominated Forests of Nepal, Central Himalaya[J]. Journal of Resources and Ecology, 2025 , 16(1) : 124 -131 . DOI: 10.5814/j.issn.1674-764x.2025.01.012
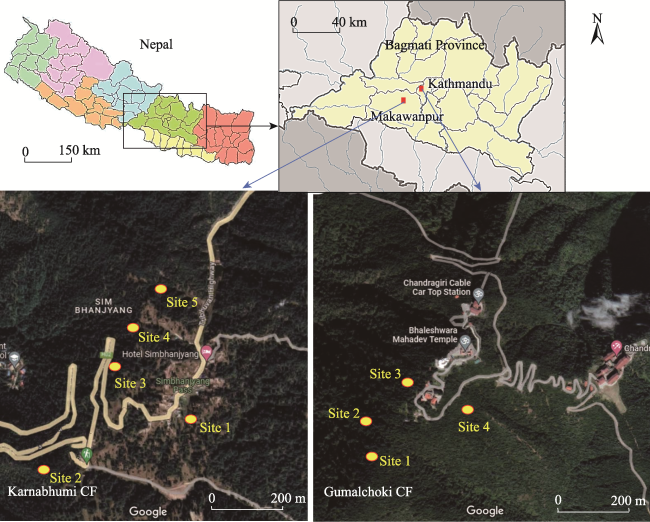
Figure 1 Study sites in Karnabhumi CF and Gumalchoki CF from Makawanpur and Kathmandu districts of Bagmati Province, central Nepal |
Table 1 Locations and bracket fungi samples collection time in two research sites in mid hills of Nepal |
| S.N. | Research sites | Latitude/Longitude | Altitude (m) | Collection year |
|---|---|---|---|---|
| 1. | Gumalchoki Community Forest (GCF), Chandragiri, Kathmandu District | 27°39'0.3''N/85°15'35.7''E | 2530 | 2017, 2020, 2021 |
| 2. | Karnabhumi Community Forest (KCF), Simbhanjyang, Makawanpur District | 27°35'41.9''N/85°04'56.0''E | 2500 | 2018, 2021, 2022 |
Table 2 Diversity, distribution and details of bracket fungi in Oak dominated forests of Nepal, central Himalaya. |
| S.N. | Species of bracket fungi | Sites | Habitat | Ecology | Edibility | Common/Local name |
|---|---|---|---|---|---|---|
| Order: Hymenochaetales Family: Hymenochaetaceae | ||||||
| 1 | Coltricia cinnamomea (Jacq.) Murrill | GCF, KCF | Fallen branches | Mycorrhizal/ Saprobic | Inedible | Cinnamon cap |
| 2 | Coltricia perennis (L.) Murrill | GCF | Soil | Saprobic | Inedible | Tiger’s eye |
| 3 | Hymenochaete rubiginosa (Dicks.) Lév. | GCF, KCF | Dead trunk | Saprobic | Inedible | Oak curtain crust |
| 4 | Inonotus hispidus (Bull.) P. Karst. | GCF, KCF | Living trunk | Parasitic | Inedible | Shaggy bracket |
| 5 | Phellinus igniarius (L.) Quél | GCF | Dead/Living trunk | Saprobic/Parasitic | Inedible | Willow bracket |
| Order: Polyporales Family: Fomitopsidaceae | ||||||
| 6 | Daedalea quercina (L.) Pers. | GCF | Dead trunk | Saprobic | Inedible | Oak mazegill |
| 7 | Fomitopsis betulina (Bull.) B.K.Cui, M.L.Han & Y.C.Dai | KCF | Living trunk | Parasitic | Inedible | Birch polypore |
| 8 | Fomitopsis pinicola (Sw) P. Karst. | GCF, KCF | Dead trunk | Saprobic | Inedible | Red-banded polypore |
| 9 | Laetiporus sulphureus (Bull.) Murrill | KCF | Dead/Living trunk | Saprobic/Parasitic | Edible/Medicinal | Sulphur polypore/ Rato chyau |
| 10 | Phaeolus schweinitzii (Fr.) Pat. | GCF | Living trunk | Parasitic | Inedible | Dyer’s mazegill |
| Family: Ganodermataceae | ||||||
| 11 | Ganoderma lucidum (Curtis) P. Karst. | GCF, KCF | Dead/Living trunk | Saprobic/Parasitic | Edible/Medicinal | Lacquered bracket/Rato chyau |
| 12 | Ganoderma sessile Murrill | KCF | Dead/Living trunk | Saprobic/Parasitic | Inedible | -- |
| Family: Irpicaceae | ||||||
| 13 | Byssomerulius corium (Pers.) Parmasto | KCF | Fallen branches | Saprobic | Unknown | -- |
| Family: Meruliaceae | ||||||
| 14 | Phlebia tremellosa (Schrad.) Nakasone & Burds | GCF, KCF | Fallen branches | Saprobic | Inedible | Jelly rot |
| Family: Polyporaceae | ||||||
| 15 | Daedaleopsis confragosa (Bolton) J. Schröt. | GCF, KCF | Living trunk | Parasitic | Inedible | Blushing bracket |
| 16 | Microporus affinis (Blume & T. Nees) Kuntze | GCF, KCF | Fallen branches | Saprobic | Unknown | -- |
| 17 | Microporus xanthopus (Fr.) Kuntze | GCF, KCF | Fallen branches | Saprobic | Unknown | Yellow-stemmed micropore |
| 18 | Pycnoporus cinnabarinus (Jacq.) P. Karst | GCF, KCF | Fallen branches | Saprobic | Inedible | Cinnabar bracket |
| 19 | Trametes elegans (Spreng.) Fr. | GCF | Fallen branches | Saprobic | Inedible | White maze polypore |
| 20 | Trametes gibbosa (Pers.) Fr. | KCF | Fallen branches | Saprobic | Inedible | Lumpy bracket |
| 21 | Trametes pubescens (Schumach.) Pilát | KCF | Fallen branches | Saprobic | Inedible | -- |
| 22 | Trametes versicolor (L.) Lloyd. | GCF, KCF | Fallen branches | Saprobic | Inedible | Turkey-tail |
| 23 | Tyromyces chioneus (Fr.) P.Karst. | KCF | Fallen branches | Saprobic | Inedible | White cheese polypore |
| Family: Steccherinaceae | ||||||
| 24 | Flabellophora superposita (Berk.) G.Cunn. | KCF | Fallen branches | Saprobic | Inedible | -- |
| Order: Russulales Family: Stereaceae | ||||||
| 25 | Stereum hirsutum (Willd.) Pers | GCF, KCF | Dead/Living trunk | Saprobic/Parasitic | Inedible | Hairy curtain crust |
| 26 | Stereum rugosum Pers. | GCF, KCF | Dead/Living trunk | Saprobic/Parasitic | Unknown | Bleeding broadleaf crust |
Figure 2 Photographs of bracket fungi reported in oak dominated forests of Nepal, central HimalayaNote: 1. Byssomerulius corium; 2. Coltricia cinnamomea; 3. Coltricia perennis; 4. Daedaleopsis confragosa; 5. Flabellophora superposita; 6. Ganoderma lucidum; 7. Ganoderma sessile; 8. Fomitopsis betulina; 9. Hymenochaete rubiginosa; 10. Microporus affinis; 11. Microporus xanthopus; 12. Phaeolus schweinitzii; 13. Phellinus igniarius; 14. Phlebia tremellosa; 15. Pycnoporus cinnabarinus; 16. Stereum hirsutum; 17. Stereum rugosum; 18. Trametes elegans; 19. Trametes gibbosa; 20. Trametes pubescens; 21. Trametes versicolor; 22. Tyromyces chioneus; 23. Daedalea quercina; 24. Fomitopsis pinicola; 25. Laetiporus sulphureus; 26. Inonotus hispidus. |
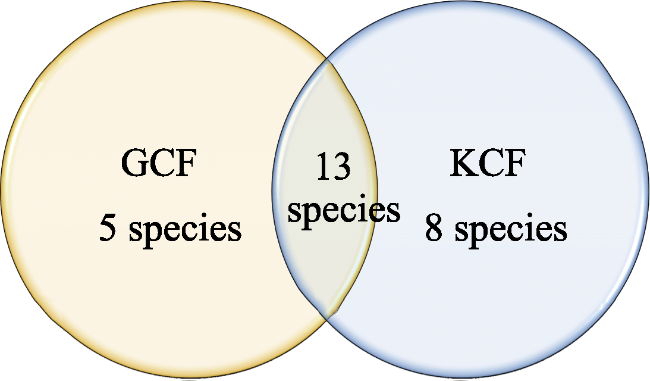
Figure 3 Venn diagram showing the number of bracket fungi in two oak dominated forests (GCF and KCF) of Nepal, central Himalaya |
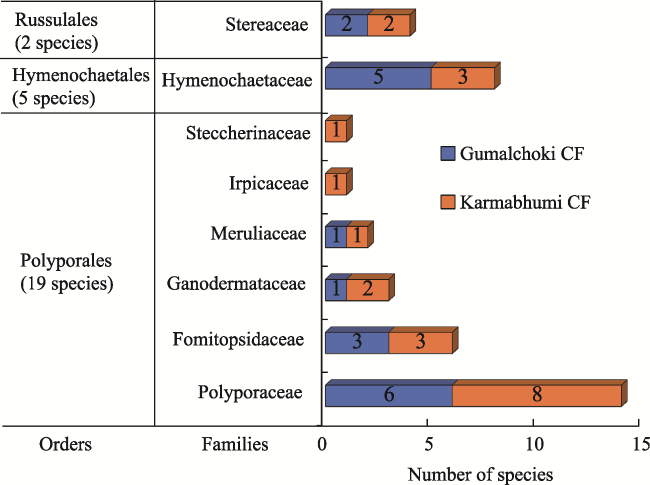
Figure 4 Diversity of bracket fungi orders in Oak dominated Forest Nepal, central HimalayaNote: Number of species in the Orders represents the overall species number of bracket fungi in both Oak dominated forests. |
Table 3 Macrofungal taxa, species richness, diversity indices, edibility, ecological role and habitat diversity of bracket fungi species in oak forest of different community forest, central Nepal |
| Variable | GCF | KCF |
|---|---|---|
| Diversity indices | ||
| Species richness | 18 | 21 |
| Shannon-Wiener Index (H) | 0.26 | 0.17 |
| Simpson Index of Diversity (1-D) | 0.53 | 0.35 |
| Margalef’s Index (Dm) | 5.22 | 6.12 |
| Pielou Index (E) | 0.078 | 0.053 |
| Edibility | ||
| Inedible | 16 | 15 |
| Unknown | 1 | 4 |
| Edible/Medicinal | 1 | 2 |
| Ecology | ||
| Saprobic | 11 | 12 |
| Saprobic/Parasitic | 3 | 5 |
| Parasitic | 4 | 3 |
| Mycorrhizal/Saprobic | 0 | 1 |
| Habitat | ||
| Fallen branches | 7 | 11 |
| Living trunk | 3 | 3 |
| Dead trunk | 3 | 2 |
| Dead/Living trunk | 4 | 5 |
| Soil | 1 | 0 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}