
Journal of Resources and Ecology >
Differences in Seedling and Sapling Densities and Species Composition between Canopy Gaps and Forest Understories in a Subtropical Forest in Bangladesh
Received date: 2024-02-05
Accepted date: 2024-07-02
Online published: 2025-01-21
Canopy openings have a vital role in forest structure, regeneration, and plant composition. In this study, we investigated and compared the species composition and densities of seedlings and saplings between canopy gaps (openings) and forest understories (in dense canopies) in a sub-tropical forest of Bangladesh. We objectively identified 42 canopy openings between transect lines and 42 main plots in dense canopies, sampled for regeneration and young tree patterns. For the regeneration study, we placed 2 m×2 m four subplots in each canopy opening and the main plot of dense canopy, thus making a total of 336 subplots. The species diversity of seedlings, saplings, and trees were significantly (P≤0.05) higher in the dense canopy than in the canopy opening. Although, most dominant and frequent regenerating species such as Diospyros pilosula, Garcinia cowa, Brownlowia elata, and Lithocarpus polystachya were common in both dense canopy and openings, the canopy openings had the highest Importance Value Index, suggesting the significance of gap dynamics in the dominance of native floral species. The dense canopy played an important role in forest compositions because 12 regenerating species, including Stereospermum suaveolens and Diospyros montana, were not found in the canopy openings, implying that rain forests must maintain a high canopy cover for regeneration.
Key words: canopy opening; disturbance; forest dynamics; native flora; seedling; sapling
Tarit Kumar BAUL , Anwarul Islam CHOWDHURY , Md Jamal UDDIN , Mohammad Kamrul HASAN , Tapan Kumar NATH , Lars Holger SCHMIDT . Differences in Seedling and Sapling Densities and Species Composition between Canopy Gaps and Forest Understories in a Subtropical Forest in Bangladesh[J]. Journal of Resources and Ecology, 2025 , 16(1) : 105 -114 . DOI: 10.5814/j.issn.1674-764x.2025.01.010
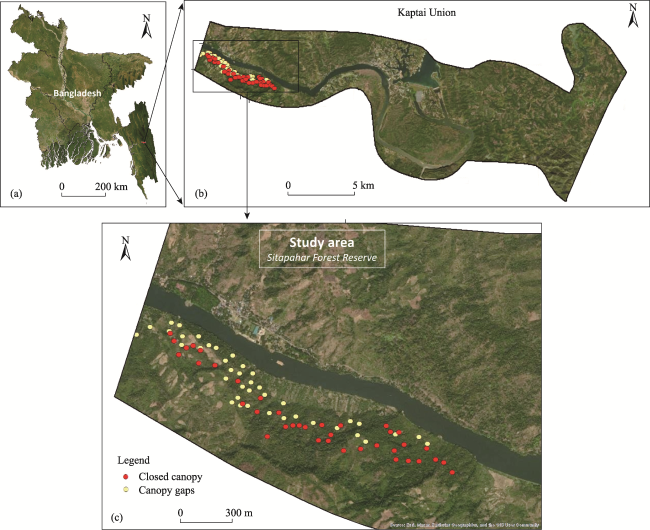
Figure 1 Sitapahar Forest Reserve (SFR) with sample plots shown (c), under Kaptai Union (b) in Bangladesh (a) |
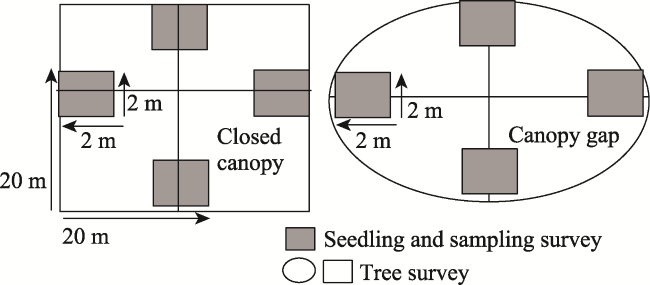
Figure 2 Layout of the sampling of trees and regenerations in dense canopy and canopy openings of SFR, Bangladesh |
Table 1 Equations used in analyses of data |
| No. | Equations for parameters | Definitions | Reference |
|---|---|---|---|
| 1 | $~~~d=\frac{\Sigma n}{\Sigma {{A}_{g}}}\times 10000$ | d: density of regenerations or trees (no. ha-1), ΣAg: total area of all openings or dense canopies, Σn: total number of individuals or regenerations of all species in each gap or dense canopies | Shukla and Chandel, 2000 |
| 2 | $H=\mathop{\sum }^{}{{p}_{i}}\times \ln \left( {{p}_{i}} \right)$ | H: Shannon-Wiener diversity index or number of species per subplot, pi: S/Σn, where S is the number of regenerations of each species and Σn is the total number of regenerations of all species | Michael, 1990 |
| 3 | $RD~=\frac{S}{\Sigma n}\times 100%$ | RD: relative density of a species (%), S: the number of regenerations of each species, Σn: total number of regenerations of all species | Misra, 1968 |
| 4 | $F=\frac{a}{b}$ | F: frequency of occurrence of a species, a: number of subplots in which regenerating species occurs, b: total number of subplots studied | Shukla and Chandel, 2000 |
| 5 | $\text{ }\!\!~\!\!\text{ }RF~=\frac{{{F}_{i}}}{\Sigma {{F}_{i}}}\times 100%$ | RF: relative frequency of a species (%), Fi: frequency of a species, ΣFi: frequencies of all species | Mori et al., 1983 |
| 6 | $A=\frac{S}{a}$ | A: abundance of a species, S: the number of regenerations of each species, a: number of subplots in which regenerating species occurs | Shukla and Chandel, 2000 |
| 7 | $RA~=\frac{{{A}_{i}}}{\Sigma {{A}_{i}}}\times 100%$ | RA: relative abundance of a species, Ai: abundance of a species, ΣAi: abundance of all species. | Misra, 1968 |
| 8 | IVI = (RD+RF+RA)×100% | IVI: Importance value index |
Table 2 Abundance, density, diversity (number of species per subplot), and mean height and collar diameter of regenerations in canopy openings and dense canopy of SFR, Bangladesh |
| Variables | Seedling | Sapling | ||
|---|---|---|---|---|
| Canopy openings (n = 168) | Dense canopy (n = 168) | Canopy openings (n = 168) | Dense canopy (n = 168) | |
| Abundance of regenerating species | 54 | 65 | 39 | 50 |
| Regenerating species | 2.63 ± 0.10 b | 4.54 ± 0.19 a | 0.95 ± 0.07 b | 2.21 ± 0.20 a |
| Regenerating individuals (density subplot-1) | 3.46 ± 0.13 b | 5.73 ± 0.25 a | 1.16 ± 0.10 b | 2.56 ± 0.23 a |
| Regeneration density (no. ha-1) | 8640.63± 333.86 b | 14312.5 ± 627.33 a | 2867.23 ± 246.38 b | 6406.25 ± 574.45 a |
| Mean height (cm) | 50.13 ± 1.61 a | 52.09 ± 1.68 a | 82.48 ± 5.44 b | 127.51 ± 6.51 a |
| Mean collar diameter (cm) | 0.69± 0.03 a | 0.76 ± 0.04 a | 0.97± 0.68 b | 1.42 ± 0.07 a |
| Number of species per subplot | 0.84 ± 0.04 b | 1.37 ± 0.05 a | 0.19 ± 0.03 b | 0.64 ± 0.07 a |
Note: Values followed by ± are standard error of mean. The same suffix letter indicates no significant difference (P>0.05) between canopy openings and dense canopies for the variables of seedlings and for saplings. |
Table 3 Relative density (RD), relative frequency (RF), relative abundance (RA), and importance value index (IVI) of seedlings of dominant species in canopy openings and dense canopy of SFR, Bangladesh |
| Seedling species | RD (%) | RF (%) | RA (%) | IVI (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Canopy openings | Dense canopy | Canopy openings | Dense canopy | Canopy openings | Dense canopy | Canopy Openings | Dense canopy | |
| Brownlowia elata | 4.7 | 2.4 | 4.8 | 2.8 | 1.9 | 1.4 | 11.4 | 6.6 |
| Diospyros pilosula | 10.3 | 7.3 | 8.3 | 6.9 | 2.4 | 1.7 | 21.0 | 15.8 |
| Ficus auriculata | 4.5 | 0.9 | 3.3 | 1.1 | 2.6 | 1.3 | 10.5 | 3.2 |
| Garcinia cowa | 8.5 | 9.9 | 7.4 | 7.5 | 2.2 | 2.1 | 18.1 | 19.5 |
| Lithocarpus acuminata | 5.4 | 3.1 | 5.7 | 3.0 | 1.8 | 1.6 | 13.0 | 7.7 |
| Lithocarpus polystachya | 10.5 | 6.6 | 11.7 | 5.0 | 1.7 | 2.1 | 23.9 | 13.7 |
| Myristica linifolia | 4.5 | 2.9 | 4.5 | 3.6 | 1.9 | 1.3 | 11.0 | 7.7 |
| Protium serratum | 1.4 | 4.6 | 1.4 | 5.0 | 1.9 | 1.5 | 4.8 | 11.1 |
| Quercus gomeziana | 5.1 | 4.2 | 5.0 | 5.0 | 1.9 | 1.3 | 12.0 | 10.5 |
| Unknown or unidentified | 4.2 | 0.9 | 4.8 | 0.8 | 1.7 | 1.7 | 10.6 | 3.4 |
Table 4 Relative density (RD), relative frequency (RF), relative abundance (RA) and importance value index (IVI) of saplings of dominant species in canopy openings and dense canopy of SFR, Bangladesh |
| Sapling species | RD (%) | RF (%) | RA (%) | IVI (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Canopy openings | Dense canopy | Canopy openings | Dense canopy | Canopy openings | Dense canopy | Canopy Openings | Dense canopy | |
| Brownlowia elata | 10.3 | 6.8 | 10.5 | 6.2 | 2.7 | 2.2 | 23.5 | 15.2 |
| Diospyros pilosula | 7.0 | 12.6 | 8.6 | 10.7 | 2.2 | 2.4 | 17.8 | 25.7 |
| Ficus auriculata | 9.7 | 1.5 | 9.9 | 1.7 | 2.7 | 1.8 | 22.3 | 4.9 |
| Garcinia cowa | 1.6 | 6.8 | 2.0 | 6.7 | 2.2 | 2.1 | 5.8 | 15.6 |
| Lithocarpus acuminata | 3.8 | 7.3 | 4.6 | 6.2 | 2.2 | 2.4 | 10.6 | 15.9 |
| Lithocarpus polystachya | 12.4 | 2.9 | 10.5 | 2.8 | 3.2 | 2.1 | 26.2 | 7.8 |
| Myristica linifolia | 7.0 | 1.5 | 5.3 | 1.1 | 3.6 | 2.6 | 15.9 | 5.2 |
| Protium serratum | 4.3 | 5.3 | 3.9 | 5.6 | 3.0 | 1.9 | 11.3 | 12.9 |
| Quercus gomeziana | 5.4 | 3.9 | 3.7 | 13.1 | ||||
| Unknown or unidentified | 5.9 | 2.9 | 4.6 | 2.2 | 3.5 | 2.6 | 14.1 | 7.8 |
Figure 3 Densities (no. ha-1) of seedling, sapling, and tree for dominant species in canopy openings ( |
Table 5 Abundance of tree species, and density, mean dbh and height of trees in canopy openings and dense canopy of SFR, Bangladesh |
| Tree variables | Canopy openings (n= 42) | Dense canopy (n= 42) |
|---|---|---|
| Abundance of species | 49 | 88 |
| Density of trees (no. ha-1) | 262.67 ± 23.22 b | 798.67 ± 140.62 a |
| Tree mean height (m) | 6.33 ± 0.06 a | 7.19 ± 0.19 a |
| Tree mean dbh (cm) | 10.01± 0.56 a | 13.52 ± 0.82 a |
Note: Values followed by ± are standard error of mean. The same suffix letter indicates no significant difference (P>0.05) between canopy openings and dense canopies for the variables of trees. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
Bangladesh National Herbarium. 2018. Vascular flora of Chittagong and the Chittagong Hill Tracts. Minisnitry of Enviornment, Forest and Climate Change, Government of the People’s Republic of Bangladesh, Dhaka, Bangladesh.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
/
| 〈 |
|
〉 |
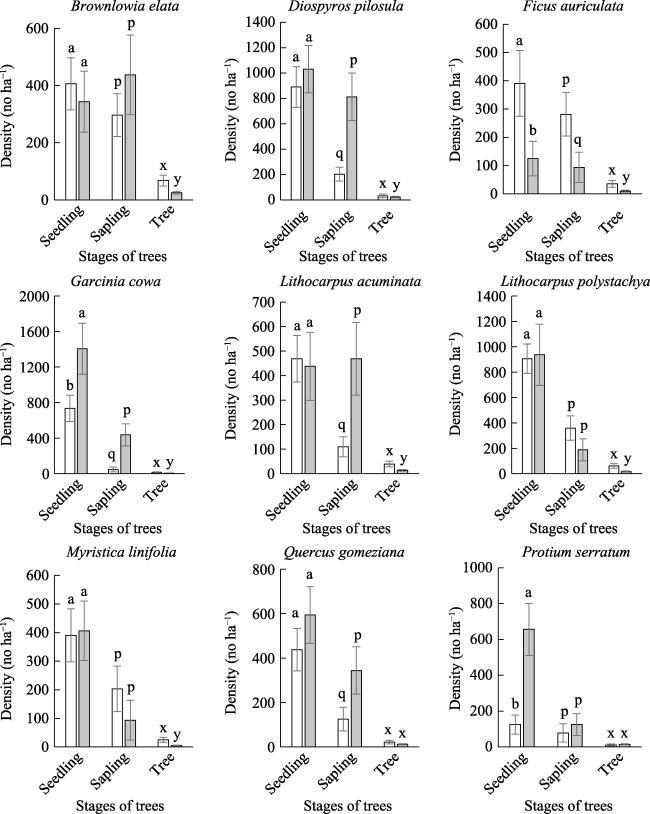
 ) and dense canopy (
) and dense canopy ( ) of SFR, Bangladesh
) of SFR, Bangladesh{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}