
Journal of Resources and Ecology >
Effect of a Cushion Plant, Androsace tapete, on Soil Net Nitrogen Mineralization and Enzyme Activities during the Growing Season
|
XING Shuo, E-mail: xings.20s@igsnrr.ac.cn |
Received date: 2022-11-20
Accepted date: 2023-05-10
Online published: 2024-03-14
Supported by
The National Natural Science Foundation of China(31770477)
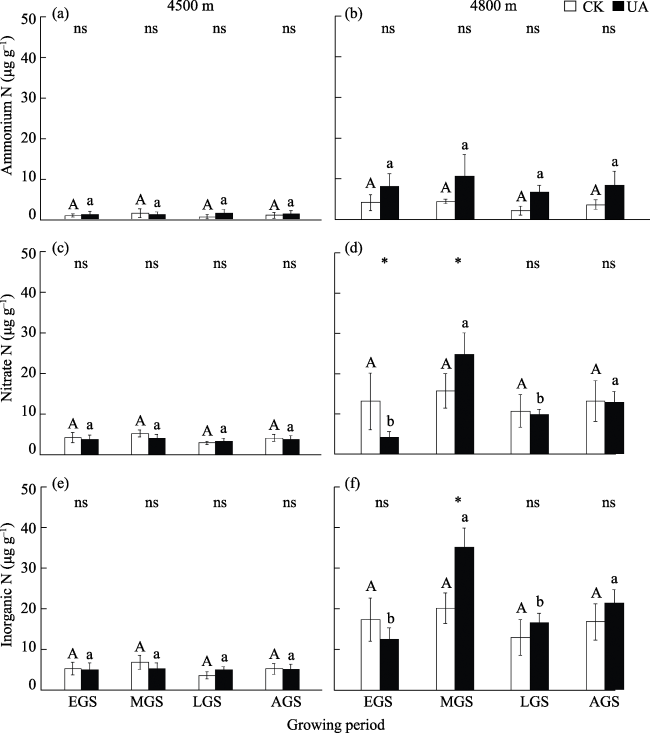
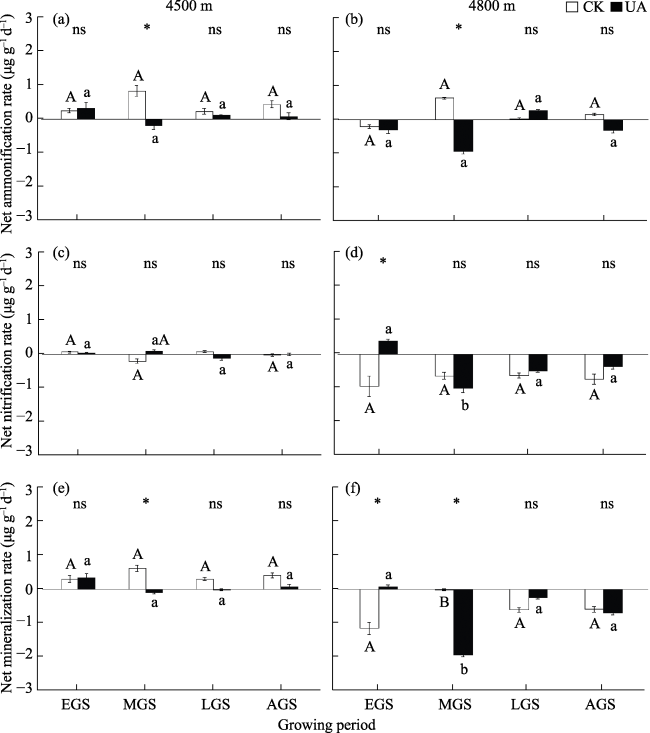
Cushion plants are one kind of unique specie in alpine ecosystems. They have a compact perennial cushion structure and play a role in facilitating the survival of other associated species by improving the local micro-environment. They are called “engineers” in the alpine ecosystem and their enhancement of soil nutrient availability is one of the ways of their engineering effect. In this study, Androsace tapete, a species of cushion plant widely distributed on the Qinghai-Tibet Plateau, was selected to investigate the dynamics of this process during the growing season at elevations of 4500 m and 4800 m on the southern slope of the Nyenchenthanglha mountains in Damxung. The effects of A. tapete on soil nutrient availability were analyzed by comparing the inorganic nitrogen content, net nitrogen mineralization rate and soil enzyme activities during the growing season in the soil under A. tapete and ambient grassland (CK). The results showed three important aspects of this system. (1) Soil nitrate nitrogen and ammonium nitrogen did not show significant differences at 4500 m, but the contents of nitrate nitrogen and inorganic nitrogen under the A. tapete soil significantly increased in the middle of the growing season at 4800 m, with nitrate nitrogen increasing by 56% and inorganic nitrogen increasing by 74.5%. (2) The trend and rate of soil nitrogen mineralization were both changed under A. tapete. In the 4500 m sample site, soil net nitrogen mineralization under A. tapete was negative (nitrogen immobilization) in the middle of the growing season, and the rate was -0.11 μg g-1 d-1, while that of CK was positive (nitrogen mineralization) and the rate was 0.61 μg g-1 d-1; and the difference between them was significant. However, both were positive in the early and late growing season, and the difference did not reach a significant level. In the 4800 m sample site, the soil net nitrogen mineralization under A. tapete was positive in the early part of the growing season, and the rate was 0.07 μg g-1 d-1, while the mineralization for CK was negative, and the rate was -1.17 μg g-1 d-1; and the difference was significant. In the middle of the growing season, both of them were negative, but the soil net nitrogen mineralization rate under A. tapete (-1.95 μg g-1 d-1) was significantly lower than that of CK (-0.02 μg g-1 d-1). At the late stage of the growing season, both of them were negative, and the difference was not significant. (3) Activities of nitrate reductase and nitrite reductase in the soil were significantly increased under A. tapete. Compared to CK at the 4500 m sample site, the activities of nitrate reductase and nitrite reductase under A. tapete were increased by 9.1% and 15.7%, respectively; and they were increased by 22.5% and 16.1%, respectively, at the 4800 m sample site. The activities of these two enzymes were significantly correlated with the dynamics of inorganic nitrogen in the soil. These results indicated that, compared to CK, the cushion plant A. tapete can change the process of soil nitrogen mineralization and the content of inorganic nitrogen, but that change had seasonal dynamics and spatial differences, which implies that this process was affected not only by the engineering of the cushion plant but also by changes in the local environment. Therefore, the engineering effect of cushion plant A. tapete was not constant during the growing seasons, and further studies are needed to clarify this process, especially considering the rapidly changing climate on the Qinghai-Tibet Plateau.
XING Shuo , HE Yongtao , NIU Ben , XU Xingliang , SONG Qian , WANG Yingfan . Effect of a Cushion Plant, Androsace tapete, on Soil Net Nitrogen Mineralization and Enzyme Activities during the Growing Season[J]. Journal of Resources and Ecology, 2024 , 15(2) : 422 -430 . DOI: 10.5814/j.issn.1674-764x.2024.02.016
Table 1 Basic physical and chemical properties of surface soil (0-10 cm) in the experimental areas |
| Elevation (m) | pH | Organic carbon (g kg-1) | Total N (g kg-1) | Bulk density (g cm-3) | C/N |
|---|---|---|---|---|---|
| 4500 | 7.03±0.22 | 29.71±4.12 | 2.65±0.39 | 1.05±0.10 | 11.69±0.10 |
| 4800 | 6.47±0.25 | 92.13±11.34 | 6.37±0.71 | 0.85±0.03 | 16.64±0.10 |
Net nitrogen mineralization=After incubation (NH4++NO3-)-Before incubation (NH4++NO3-)
Table 2 Soil water contents under A. tapete and CK |
| Sampling date | Soil sampling plot | |||
|---|---|---|---|---|
| 4500 m | 4800 m | |||
| Under A. tapete (%) | CK(%) | Under A. tapete (%) | CK(%) | |
| 2021-05-18 | 8.04±2.19a | 9.20±0.99a | 10.48±1.76b | 16.54±5.76a |
| 2021-07-21 | 20.37±2.71a | 22.96±3.33a | 48.46±8.81a | 36.56±5.41b |
| 2021-09-06 | 25.24±2.98a | 25.69±1.40a | 43.42±3.18a | 35.98±9.72b |
Note: Different letters indicate significant differences under A. tapete and CK. |
Fig. 1 Dynamics of soil inorganic nitrogen under A. tapete and CK during the growing seasonNote: CK: Without A. tapete; UA: Under A. tapete; EGS: Early growing season; MGS: Middle growing season; LGS: Late growing season; AGS: Average value of growing season. Different capital letters indicate the soil inorganic nitrogen content of CK is significantly different in the different growing periods (P<0.05). Different lowercase letters indicate the soil inorganic nitrogen content under A. tapete is significantly different in the different growing periods (P<0.05); ns: P>0.05; * indicates a significant difference between UA and CK (P<0.05). The same notations are used in subsequent figures. |
Fig. 2 Soil net nitrogen mineralization rates under A. tapete and CK during the growing season |
Table 3 Soil enzyme activities under A. tapete and CK |
| Soil enzyme | Soil sampling plot | |||
|---|---|---|---|---|
| 4500 m | 4800 m | |||
| Under A. tapete | CK | Under A. tapete | CK | |
| Urease | 52.30±9.83a | 50.08±9.47a | 81.57±18.91a | 84.23±9.47a |
| Protease | 14.85±1.34a | 14.39±1.06a | 16.78±1.66a | 16.15±1.65a |
| Nitrate reductase | 100.23±7.39a | 91.86±7.75b | 121.05±16.18a | 98.83±16.87b |
| Nitrite reductase | 43.28±12.73a | 37.40±13.34b | 25.15±3.96a | 21.66±6.26b |
Note: Different letters indicate significant differences under A. tapete and CK. |
Table 4 Correlations between soil enzyme activities and soil net nitrogen mineralization |
| Type | Nitrate nitrogen | Ammonium nitrogen | Inorganic nitrogen | Soil water content |
|---|---|---|---|---|
| Urease | 0.015 | 0.102 | 0.156 | 0.009 |
| Protease | 0.173 | 0.164 | 0.168 | 0.073 |
| Nitrate reductase | 0.480** | 0.348* | 0.451** | 0.019 |
| Nitrite reductase | 0.224 | 0.413** | 0.309* | 0.025 |
Note: **P<0.01 indicates the difference is extremely significant; *P<0.05 indicates the difference is significant. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}