
Journal of Resources and Ecology >
Characteristics of Water Consumption of Atraphaxis bracteata A. Los. in the Mu Us Sandy Land of North Central China
|
LI Wanying, E-mail: 929981653@qq.com |
Received date: 2022-06-05
Accepted date: 2023-02-20
Online published: 2023-07-14
Supported by
Key Research and Development Program of China(2017YFC0504403)
Water consumption by Atraphaxis bracteata A. Los. was measured using sap flow measuring system from May to October in 2009, and environmental variables were measured simultaneously in this study,. The study revealed the sap flow rate was largest in June, and the daily sap flow was significantly affected by weather condition. The sap flow rate is related to stem diameter, stems with larger diameter had higher sap flow rate than smaller diameter stems. The effects of soil moisture at depths of <1 m on the sap flow rate were not significant, which may be related to the uptake of water by plants from deep soil. In addition, time lags of 30-130 minutes were observed between transpiration and increased sap flow. These time lags were related to stem size as well as water conditions. The ratio of nocturnal to diurnal sap flow normally varied in the range between 0.1 and 0.4; however, on rainy days, the ratio was even larger than 1.0, indicating that nocturnal sap flow results in significant water loss. Environment factors had significant effect on sap flow. Solar radiation and the vapor-pressure deficit had the largest effect on the sap flow rate during the day and at night, respectively.
Key words: environmental factors; sap flow; soil moisture; time lag
LI Wanying , GUO Yue , WU Rina , CAO Qiqi , DING Guodong , XIAO Huijie . Characteristics of Water Consumption of Atraphaxis bracteata A. Los. in the Mu Us Sandy Land of North Central China[J]. Journal of Resources and Ecology, 2023 , 14(4) : 880 -892 . DOI: 10.5814/j.issn.1674-764x.2023.04.020
Fig. 1 The location of Yanchi research station in China |
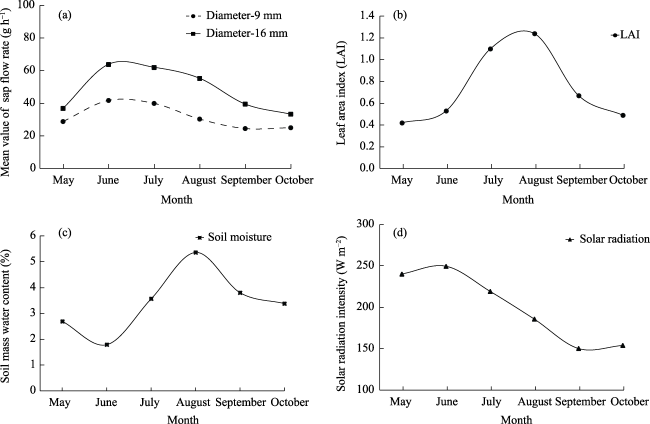
Fig. 2 Monthly variation of (a) sap flow rate of 9- and 16-mm stems of A. bracteata, (b) leaf area index of 16-mm stems of A. bracteata, (c) soil water content, (d) solar radiation intensity from May to October in 2009 |
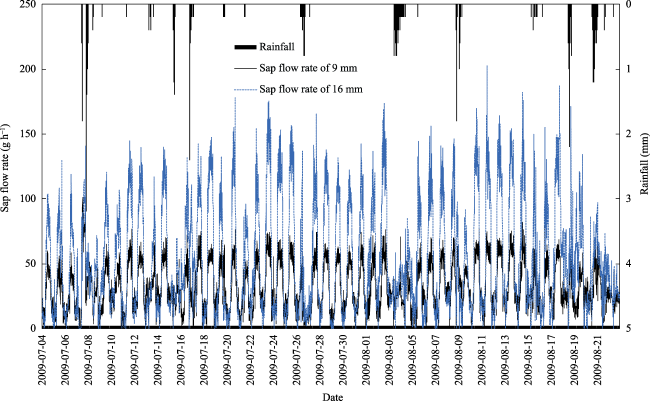
Fig. 3 Variations in sap flow rate in A. bracteata during July and August 2009 |
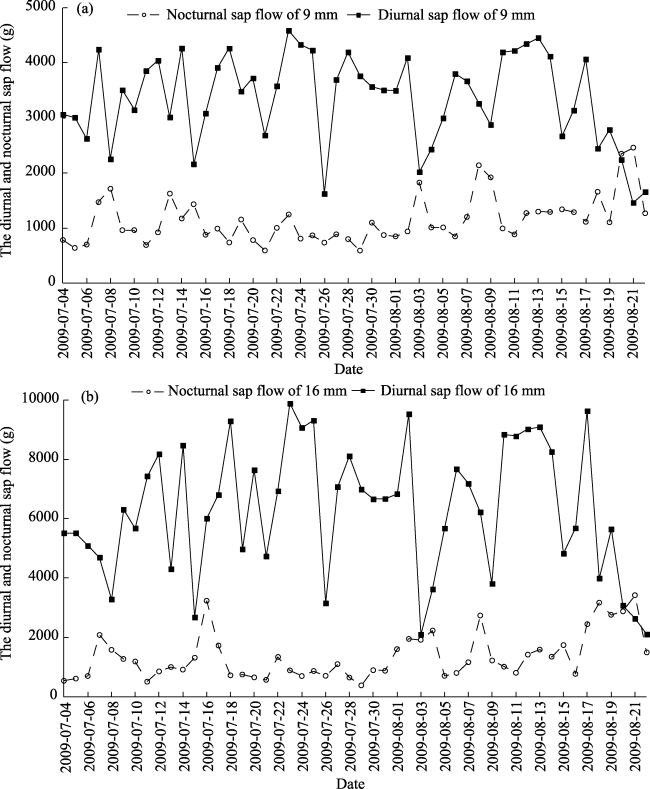
Fig. 4 Diurnal and nocturnal sap flow ratesin A. bracteata during July and August 2009 |
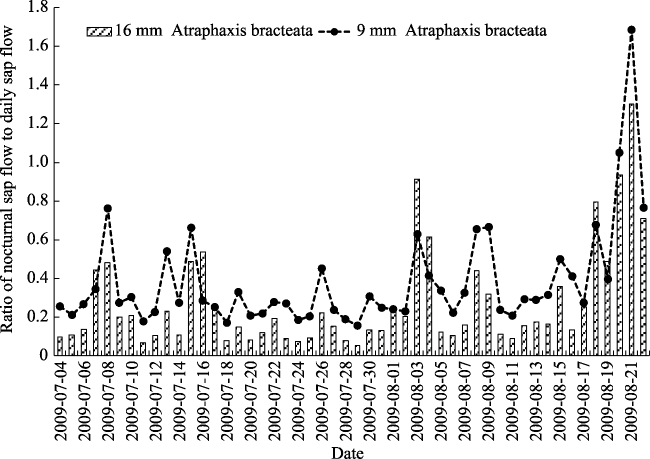
Fig. 5 Ratio of nocturnal to diurnal sap flow in A. bracteata during July and August 2009 |
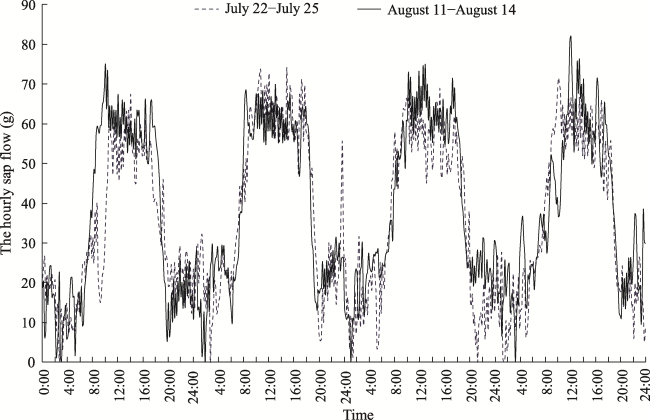
Fig. 6 Hourly sap flow of 9-mm-diameter A. bracteata stems from 22 to 25 July 2009, where soil mass water content changed from 1.67% to 2.21%; and from 11 to 14 August 2009, where soil mass water content changed from 4.98% to 4.21%Note: The horizontal axis displays four days time periods (from 0 am to 12 pm every day) in two months, from 22 to 25 July, and from 11 to 14 August. |
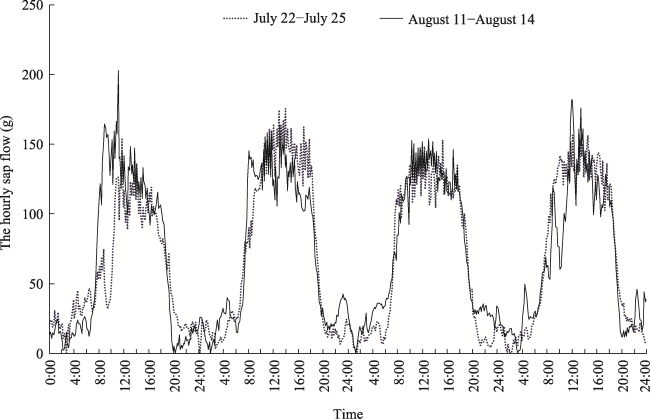
Fig. 7 Hourly sap flow of 16-mm-diameter A. bracteata from 22 to 25 July 2009, where soil mass water content changed from 1.67% to 2.21%; and from 11 to 14 August 2009, where soil mass water content changed from 4.98% to 4.21%Note: The horizontal axis displays four days time periods (from 0 am to 12 pm every day) in two months, from 22 to 25 July, and from 11 to 14 August. |
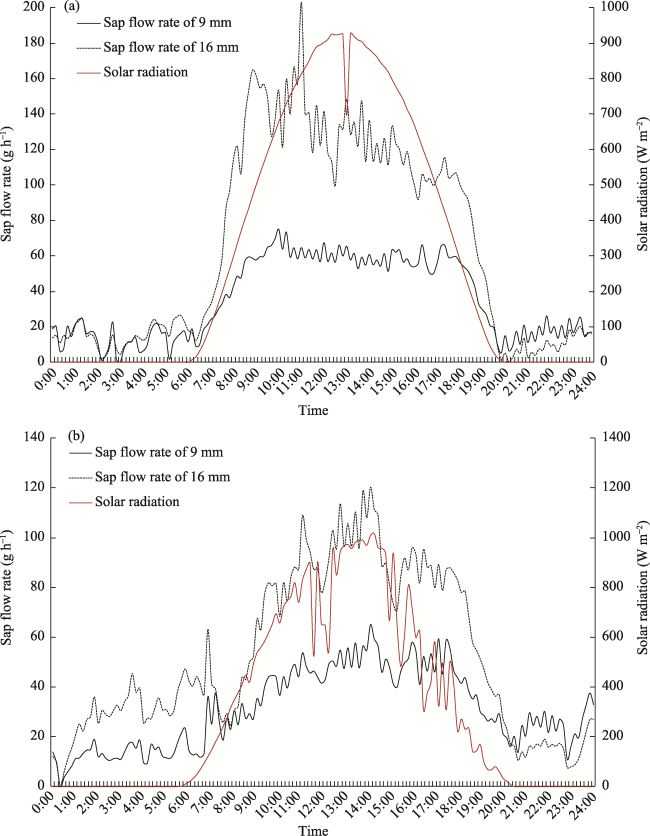
Fig. 8 Sap flow rate from 0:00 to 23:50 on 11 August (wet soil) and on 9 July (dry soil) 2009Note: The time lag between transpiration and sap flow is plotted in this figure, since the onset of transpiration occurred as solar radiation was larger than 0; however, the sap flow did not increase correspondingly. |
Table 1 VIF of the explanatory variables based on collinearity statistics |
| Explanatory variables | Collinearity statistics | |
|---|---|---|
| Tolerance | VIF | |
| Solar radiation | 0.539 | 1.854 |
| Temperature | 0.231 | 4.327 |
| Wind speed | 0.647 | 1.545 |
| Vapor pressure deficit (VPD) | 0.195 | 5.128 |
Table 2 Results of the regression model for diurnal sap flow |
| Variable | Model of 9 mm A. bracteata | Model of 16 mm A. bracteata | ||||
|---|---|---|---|---|---|---|
| Coefficient | Standardized coefficient | P | Coefficient | Standardized coefficient | P | |
| Constant | 1.295 | 0.415 | -2.709 | 0.372 | ||
| Solar radiation | 0.031 | 0.539 | <0.001 | 0.081 | 0.599 | <0.001 |
| Wind speed | 1.299 | 0.367 | <0.001 | |||
| Temperature | 0.561 | 0.059 | <0.001 | 1.956 | 0.232 | <0.001 |
| VPD | -1.174 | -0.069 | 0.012 | 4.994 | 0.123 | <0.001 |
| R2 | 0.606 | - | <0.001 | 0.735 | - | <0.001 |
Table 3 Results of the regression model for nocturnal sap flow |
| Variable | Model of 9-mm A. bracteata | Model of 16-mm A. bracteata | ||||
|---|---|---|---|---|---|---|
| Coefficient | Standardized coefficient | P | Coefficient | Standardized coefficient | P | |
| Constant | 25.296 | 0.000 | 33.419 | 0.000 | ||
| Wind speed | 1.232 | 0.158 | 0.000 | 2.090 | 0.157 | 0.000 |
| Temperature | -0.408 | -0.131 | 0.000 | -0.404 | -0.076 | 0.011 |
| VPD | -8.408 | -0.259 | 0.000 | |||
| R2 | 0.038 | - | 0.000 | 0.130 | - | 0.000 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}