
Journal of Resources and Ecology >
Diet Analysis of Asian Elephants Using Next-Generation Sequencing
|
PENG Xiaoxu, E-mail: pxx424@foxmail.com |
Received date: 2022-05-26
Accepted date: 2022-08-09
Online published: 2023-04-21
Supported by
The Pilot Study on Habitat Modification of Asian Elephants(20210126)
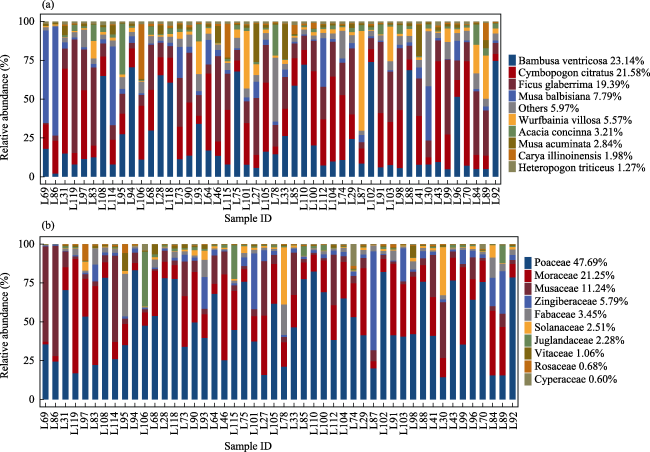
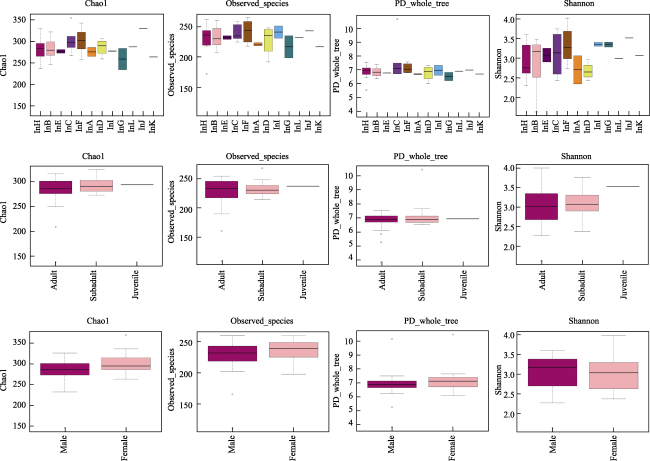
Understanding the diet composition and preferences of large herbivores not only provides insights into their ecological role, but also helps to assess the viability of elephant populations and their habitats. This study was performed to determine the diet preferences of a small population of Asian elephants in Nangunhe National Nature Reserve in Yunnan, China, during the annual dry season. The next-generation sequencing of the rbcL gene from non-invasively collected fecal samples was conducted in addition to transect surveys and camera-trapping along known elephant trails. With the transect survey, we identified 31 plant species foraged by elephants. The next-generation sequencing analysis identified a total of 90 plant species from the elephant dung samples. Only nine species were detected at rates greater than 1% in all the samples. Poaceae (47.69%), Moraceae (21.25%), and Musaceae (11.24%) were detected to have the highest rates at the family level. We also examined whether differences existed between sexes, age groups, and individuals; however, significant differences were found only between individuals. This study provides useful insights into the foraging preferences of Asian elephants, which could help in further understanding the interactions between elephants and their habitat in the reserve and inform future management decisions in related areas. The detected core plant species with relatively high abundance could provide guidance for habitat restoration and cultivation of food bases. The local plantations where the elephants prefer to feed could be moved farther away, making them inaccessible to the elephants.
PENG Xiaoxu , SUN Yakuan , CHEN Ying , Aliana NORRIS , SHI Kun . Diet Analysis of Asian Elephants Using Next-Generation Sequencing[J]. Journal of Resources and Ecology, 2023 , 14(3) : 616 -630 . DOI: 10.5814/j.issn.1674-764x.2023.03.016
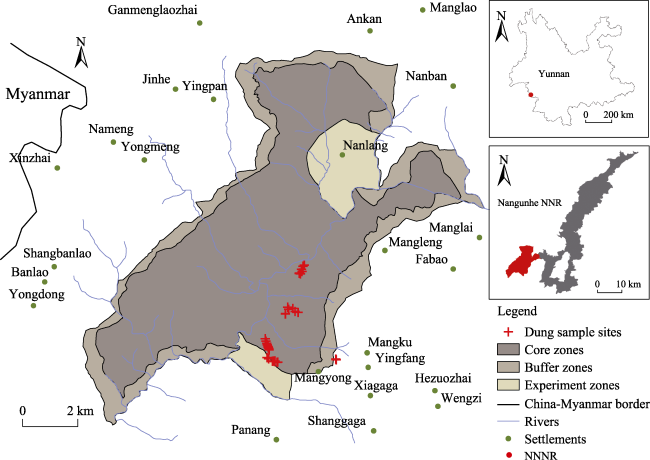
Fig. 1 Nangunhe National Nature Reserve (NNNR) map with dung sample sitesNote: Camera sites refer to Sun et al. (2021). |
Table 1 Demographic information of Asian elephant population in NNNR (Chen et al., 2022b) |
| Individual | Sex | Age |
|---|---|---|
| InA | Female | Subadult |
| InB | Female | Adult |
| InC | Female | Subadult |
| InD | Female | Adult |
| InE | Male | Subadult |
| InF | Female | Adult |
| InG | Male | Adult |
| InH | Male | Adult |
| InI | Male | Subadult |
| InJ | Male | Juvenile |
| InK | Male | Subadult |
| InL | Male | Subadult |
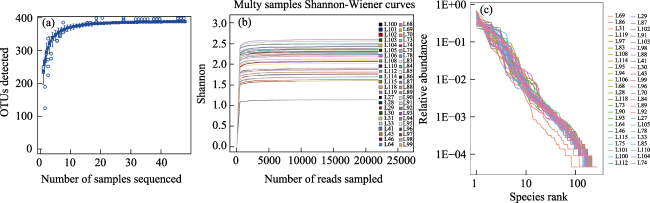
Fig. 2 Overall description of NGS sequencing resultsNote: (a) Species accumulation curve; (b) Shannon-Wiener curve, indicating the sample size and sampled reads are sufficient for subsequent analysis; (c) Rank abundance curve identifying abundant species. |
Table 2 Taxonomic classification of 117 plant species foraged by Asian elephant in NNNR |
| Categories | Family | Genus | Species | Method |
|---|---|---|---|---|
| Equisetophyta | ||||
| Equisetopsida | ||||
| Poales | Poaceae | Bambusa | Bambusa ventricosa McClure * | NGS |
| Bambusa vulgaris Schrader ex Wendland 'Wamin' * | NGS | |||
| Cymbopogon | Cymbopogon citratus (DC.) Stapf * | NGS | ||
| Heteropogon | Heteropogon triticeus (R. Br.) Stapf ex Craib. * | NGS | ||
| Panicum | Panicum incomtum Trin. * | NGS | ||
| Panicum miliaceum L. * | NGS | |||
| Oryza | Oryza rufipogon Griff. * | NGS | ||
| Oryza sativa L. | NGS | |||
| Thysanolaena | Thysanolaena latifolia (Roxb. ex Hornem.) Honda | NGS | ||
| Setaria | Setaria palmifolia (Koen.) Stapf | NGS | ||
| Setaria faberi R.A.W.Herrm. * | NGS | |||
| Dichanthium | Dichanthium annulatum (Forssk.) Stapf * | NGS | ||
| Dendrocalamus | Dendrocalamus giganteus Munro | NGS | ||
| Dendrocalamus membranaceus Munro | TS | |||
| Dendrocalamus semiscandens Hsueh et D. Z. Li * | TS | |||
| Triticum | Triticum aestivum L. | NGS | ||
| Chrysopogon | Chrysopogon zizanioides (L.) Roberty * | NGS | ||
| Phragmites | Phragmites australis (Cav.) Trin. ex Steud. | NGS | ||
| Garnotia | Garnotia tenella (Arn. ex Miq.) Janowski * | NGS | ||
| Zea | Zea mays L. | NGS | ||
| Microstegium | Microstegium vimineum (Trin.) A. Camus * | TS | ||
| Cyperaceae | Scleria | Scleria parvula Steud. * | NGS | |
| Machaerina | Machaerina rubiginosa (Biehler) T.Koyama * | NGS | ||
| Magnoliopsida | ||||
| Rosales | Moraceae | Ficus | Ficus glaberrima Bl. * | NGS |
| Ficus tinctoria Forst. | NGS | |||
| Ficus hirta Vahl | NGS | |||
| Ficus chapaensis Gagnep. * | TS | |||
| Ficus racemosa L. | TS | |||
| Ficus auriculata Lour. | TS | |||
| Streblus | Streblus indicus (Bur.) Corner * | NGS | ||
| Broussonetia | Broussonetia papyrifera (Linn.) L'Hér. ex Vent. | NGS | ||
| Morus | Morus alba L. | NGS | ||
| Morus macroura Miq. * | TS | |||
| Artocarpus | Artocarpus lakoocha Roxburgh | TS | ||
| Artocarpus pithecogallus C. Y. Wu * | TS | |||
| Rosaceae | Rubus | Rubus lambertianus Ser. * | NGS | |
| Rubus pluribracteatus L.T.Lu & Boufford | NGS, TS | |||
| Rubus pungens Cambess. * | NGS | |||
| Prunus | Prunus mira Koehne * | NGS | ||
| Urticaceae | Debregeasia | Debregeasia orientalis C. J. Chen * | NGS | |
| Cannabaceae | Trema | Trema orientale (L.) Blume * | NGS | |
| Humulus | Humulus scandens (Lour.) Merr. * | NGS | ||
| Rhamnaceae | Berchemia | Berchemia floribunda (Wall.) Brongn. * | NGS | |
| Zingiberales | Musaceae | Musa | Musa balbisiana Colla * | NGS |
| Musa itinerans Cheesm. | NGS, TS | |||
| Musa acuminata Colla | NGS | |||
| Ensete | Ensete glaucum (Roxb.) Cheesm. | NGS | ||
| Sorghum | Sorghum×drummondii (Nees ex Steud.) Millsp. & Chase * | NGS | ||
| Zingiberaceae | Wurfbainia | Wurfbainia villosa (Lour.) * | NGS | |
| Alpinia | Alpinia galanga (L.) Willd. * | NGS | ||
| Marantaceae | Donax | Donax canniformis (Forst.) K. Schum. * | NGS | |
| Fabales | Fabaceae | Acacia | Acacia concinna (Willd.) DC. * | NGS |
| Acacia pennata (L.) Willd. | TS | |||
| Pueraria | Pueraria montana (Lour.) Merr. | NGS | ||
| Bauhinia | Bauhinia touranensis Gagnep. * | NGS | ||
| Bauhinia acuminate L. | TS | |||
| Senegalia | Senegalia catechu (L.f.) P.J.H.Hurter & Mabb. * | NGS | ||
| Mucuna | Mucuna pruriens (L.) DC. * | NGS | ||
| Arachis | Arachis hypogaea Linn.bam * | NGS | ||
| Albizia | Albizia crassiramea Lace * | TS | ||
| Fagales | Juglandaceae | Carya | Carya illinoiensis K. Koch * | NGS |
| Carya illinoiensis K. Koch * | NGS | |||
| Fagaceae | Castanea | Castanea seguinii Dode * | NGS | |
| Quercus | Quercus baronii Skan * | NGS | ||
| Castanopsis | Castanopsis indica (Roxburgh ex Lindley) A. DC. * | TS | ||
| Vitales | Vitaceae | Tetrastigma | Tetrastigma subtetragonum C. L. Li * | NGS |
| Tetrastigma delavayi Gagnep. * | NGS | |||
| Tetrastigma planicaule (Hook.) Gagnep. | NGS, TS | |||
| Tetrastigma tsaianum C. Y. Wu * | NGS | |||
| Leea | Leea indica (Burm. f.) Merr. | NGS | ||
| Cucurbitales | Begoniaceae | Begonia | Begonia palmata D. Don * | NGS |
| Begonia masoniana Irmsch. ex Ziesenh. * | NGS | |||
| Cucurbitaceae | Hodgsonia | Hodgsonia macrocarpa (Bl.) Cogn. * | NGS | |
| Gynostemma | Gynostemma pentaphyllum (Thunb.) Makino * | NGS | ||
| Ranunculales | Menispermaceae | Tinospora | Tinospora crispa (L.) Hook. f. et Thomson * | NGS |
| Parabaena | Parabaena sagittata Miers ex Hook.f. & Thomson | NGS | ||
| Brassicales | Brassicaceae | Arabidopsis | Arabidopsis thaliana (L.) Heynh. * | NGS |
| Icacinales | Icacinaceae | Iodes | Iodes cirrhosa Turcz. * | NGS |
| Arecales | Arecaceae | Caryota | Caryota maxima Blume * | NGS, TS |
| Caryota obtusa Griffith | TS | |||
| Salacca | Salacca zalacca (Gaertn.) Voss * | NGS | ||
| Calamus | Calamus gracilis Roxb. * | NGS | ||
| Calamus nambariensis Becc. | NGS | |||
| Laurales | Lauraceae | Cinnamomum | Cinnamomum verum Presl * | NGS |
| Cinnamomum parthenoxylon (Jack) Meisn. * | NGS | |||
| Phoebe | Phoebe puwenensis Cheng * | TS | ||
| Asparagales | Hypoxidaceae | Curculigo | Curculigo capitulata (Lour.) Ktze. | NGS |
| Orchidaceae | Thrixspermum | Thrixspermum japonicum (Miq.) Rchb. f. * | NGS | |
| Cornales | Nyssaceae | Nyssa | Nyssa yunnanensis W. C. Yin * | NGS |
| Camptotheca | Camptotheca acuminata Decne. * | NGS | ||
| Quercus | Quercus baronii Skan * | NGS | ||
| Solanales | Solanaceae | Solanum | Solanum lycopersicum L. * | NGS |
| Convolvulaceae | Ipomoea | Ipomoea sumatrana (Miq.) Ooststr. * | NGS | |
| Caryophyllales | Polygonaceae | Persicaria | Persicaria chinensis (L.) H.Gross * | NGS |
| Persicaria runcinata (Buch.-Ham. ex D.Don) H.Gross * | NGS | |||
| Liliales | Smilacaceae | Smilax | Smilax china L. * | NGS |
| Smilax ocreata A.DC. | NGS | |||
| Ericales | Theaceae | Camellia | Camellia oleifera Abel.c * | NGS |
| Apiales | Araliaceae | Trevesia | Trevesia palmata (Roxb. ex Lindl.) Vis. | NGS |
| Aralia | Aralia thomsonii Seem. | TS | ||
| Malvales | Malvaceae | Kleinhovia | Kleinhovia hospita L. * | NGS |
| Sterculia | Sterculia lanceolata Cav. * | TS | ||
| Lamiales | Orobanchaceae | Rehmannia | Rehmannia glutinosa (Gaetn.) Libosch. ex Fisch. et Mey. * | NGS |
| Lamiaceae | Gmelina | Gmelina arborea Roxb. * | TS | |
| Malpighiales | Clusiaceae | Garcinia | Garcinia xipshuanbannaensis Y. H. Li * | NGS |
| Phyllanthaceae | Aporosa | Aporosa yunnanensis (Pax & K.Hoffm.) F.P.Metcalfarabidopsis * | NGS | |
| Baccaurea | Baccaurea ramilflora Loureiro | TS | ||
| Euphorbiaceae | Mallotus | Mallotus paniculatus (Lam.) Muell. Arg. * | TS | |
| Mallotus japonicus (Thunb.) Muell. Arg. * | TS | |||
| Macaranga | Macaranga denticulata (Bl.) Muell. Arg. | TS | ||
| Homonoia | Homonoia riparia Lour. | TS | ||
| Piperales | Piperaceae | Piper | Piper sarmentosum Roxb. * | NGS |
| Sapindales | Sapindaceae | Pometia | Pometia pinnata J. R. et G. Frost. * | TS |
| Aquifoliales | Aquifoliaceae | Ilex | Ilex polypyrene C. J. Tseng et B. W. Liu * | TS |
| Proteales | Sabiaceae | Meliosma | Meliosma arnottiana Walp. * | TS |
| Pinopsida | ||||
| Gnetales | Gnetaceae | Gnetum | Gnetum pendulum C.Y.Cheng * | NGS |
| Gnetum montanum Markgr. | TS |
Note: NGS: next-generation sequencing; TS: transect survey; *: new record of Asian elephant diet; the four common species detected by NGS and transect survey are in bold font. |
Fig. 3 Stacked bar-graph showing the top 10 (a) species and (b) families by relative abundance |
Fig. 4 Box plots of alpha diversity indexNote: Individual, age, and sex groups for each row; Chao1, Observed species, PD whole tree, and Shannon index for each column. No significant differences existed for alpha diversity. |
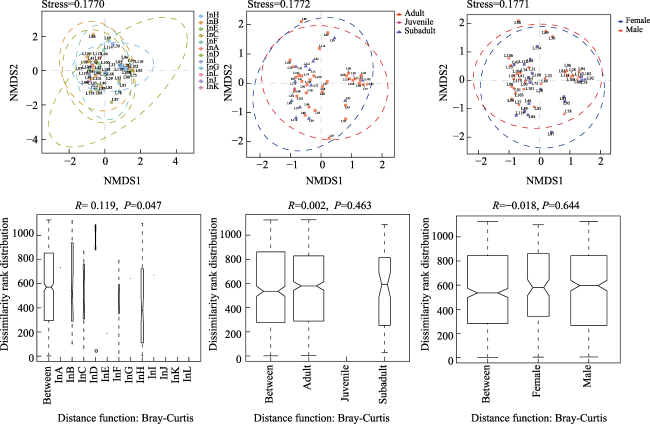
Fig. 5 NMDS plot and ANOSIM analysis of individual, age and sex groups for each column with stress < 0.2 according to Bray-Curtis distance; no significant clustering existed between age and sex (ANOSIM P > 0.05). Significant differences existed between individuals (ANOSIM P<0.05) |
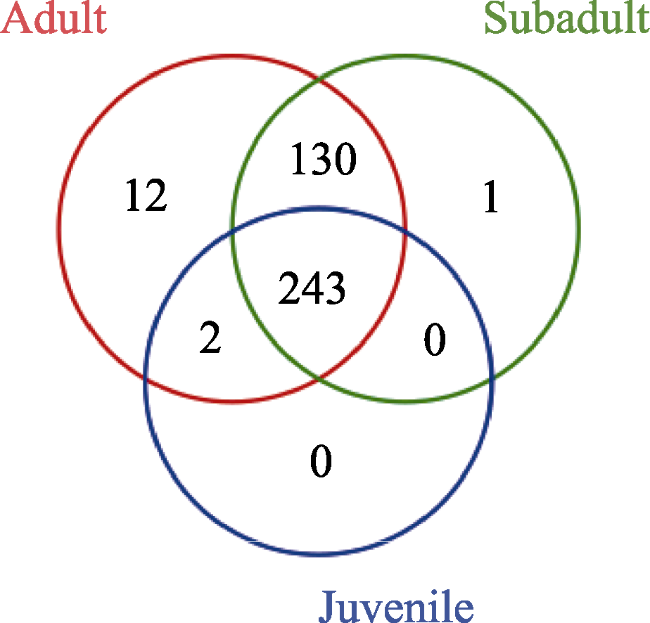
Fig. 6 Venn diagram of age groups showing number of mutual and unique OTUs |
This study was supported by the Second National Survey of Terrestrial Wildlife Program funded by the National Forestry and Grassland Administration of China. We thank WANG Jiahui, LIU Ying, and LIU Zhen who provided support in field surveys. We also acknowledge Luciano Atzeni and LI Fengjiao for their comments on the manuscript.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
CVH Chinese Virtual Herbarium. 2003. National plant specimen resource center. https://www.cvh.ac.cn/.Viewed on 2 April 2020. on 2 April 2020.
|
| [19] |
|
| [20] |
Editorial Committee of FRPS. 1993. Flora Reipublicae Popularis Sinicae. Beijing, China: Science Press. (in Chinese)
|
| [21] |
|
| [22] |
Flora of China Editorial Committee. 2013. Flora of China. Beijing, China: Science Press. (in Chinese)
|
| [23] |
FRPS Flora Reipubliae Popularis Sinicae. 2005. www.iplant.cn/frps. Viewed on 2 April 2020.
|
| [24] |
|
| [25] |
Group C P W. 2009. A DNA barcode for land plants. Proceedings of the National Academy of Sciences of USA, 106(31): 12794-12797.
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
Missouri Botanical Garden. 2012. Tropicos. https://www.tropicos.org. Viewed on 2 April 2020.
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
POWO Plants of the World Online. 2019. Plants of the world online. https://powo.science.kew.org/, Viewed on 2 April 2020.
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
Royal Botanic Garden. 2010. The plant list (version 1).http://www.theplantlist.org/. Viewed on 2 April 2020.
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}