
Journal of Resources and Ecology >
Response of Natural Regeneration of Pinus massoniana and Quercus variabilis Mixed Forest to Thinning Intensity and Environmental Factors
|
GUO Shiyu, E-mail: guoshiyu3201@163.com |
Received date: 2021-12-02
Accepted date: 2022-04-02
Online published: 2023-02-21
Supported by
The Key Forestry Research Project Sponsored by Sino-German Financial Cooperation(zdczhz2021ky09)
The Key Forestry Research Project Sponsored by Sino-German financial cooperation(zdczhz2019ky01)
The Forest Sustainable Management Project Sponsored by Sino-German Financial Cooperation in Southern China (Hubei Province)(BMZ ID 2006 65 778)
Based on target tree management, the effects of different thinning intensities and environmental factors on the natural regeneration of a Pinus massoniana and Quercus variabilis mixed forest were explored in order to provide a theoretical basis for the natural regeneration and sustainable forest management of P. massoniana and Quercus L. mixed forests. Taking the mixed forest after thinning as the research object, three average thinning intensities of WT (7.6%) for weak thinning, LT (15.3%) for light thinning, and MT (24.3%) for moderate thinning were carried out in 5 m×5 m quadrats with 7-10 replicates for each intensity level and 3 replicates for the control. Three years after the thinning, the amount of natural regeneration, growth height, regeneration density, diversity of regenerated tree species and their influencing factors at different thinning intensities were measured and analyzed. The results indicated four main features of the subsequent regeneration. (1) There were 32 species of vascular plants in the 28 quadrats 3 years after thinning, belonging to 22 families and 30 genera, and the dominant species for regeneration were arbor species. The number of regeneration species increased with increasing thinning intensity. (2) As thinning intensity increased, the number of natural regeneration plants between various height classes rose; so, the increased thinning intensity promoted the density of different height classes during regeneration. (3) As thinning intensity increased, so did species abundance S and species evenness. The degree and intrinsic diversity increased, while the Shannon-Weiner and Simpson indices showed no discernible trends. (4) Slope, aspect, and slope position, as well as thinning intensity, all had significant impacts on species richness, species evenness, and regeneration density. MT has the most appropriate promoting effect on natural regeneration and species diversity, so increased thinning intensity can promote natural regeneration and species diversity in the P. massoniana and Q. variabilis mixed forest. In addition, aspect and slope position can increase the species richness S and diversity of natural regeneration, whereas slope has a clear inhibitory effect on the species richness S and diversity during natural regeneration.
GUO Shiyu , SONG Dekai , XU Zijing , CHEN Shiyun , CHEN Zeyan , DU Peng , WANG Yang . Response of Natural Regeneration of Pinus massoniana and Quercus variabilis Mixed Forest to Thinning Intensity and Environmental Factors[J]. Journal of Resources and Ecology, 2023 , 14(2) : 423 -432 . DOI: 10.5814/j.issn.1674-764x.2023.02.020
Table 1 Basic conditions of sampling plots |
| Thinning intensity | Quadrat No. | Composition of trees | Aspect | Slope (°) | Slope position | DBH (cm) | Tree height (m) | Canopy density | Stand density after thinning (plants ha-1) |
|---|---|---|---|---|---|---|---|---|---|
| WT | 1 | Pm: Qv: Cf (7:2:1) | E | 6 | up | 13.00±3.72 | 10.67±1.22 | 0.82 | 1577 |
| 2 | Pm: Qv :Cf (6:3:1) | NW | 7.5 | middle | 13.36±5.06 | 11.04±1.21 | 0.84 | 1535 | |
| 3 | Pm: Qv: Cf: Ic (4: 4:1:1) | S | 6.5 | middle | 12.96±5.18 | 10.54±1.23 | 0.84 | 1395 | |
| 4 | Pm: Qv: Cf (6:3:1) | SE | 5 | down | 15.14±4.56 | 12.22±1.24 | 0.85 | 1231 | |
| 5 | Pm: Qv (7:3) | NW | 5 | down | 15.38±4.75 | 12.56±1.22 | 0.81 | 1382 | |
| 6 | Pm: Qv: Cf (7:2:1) | E | 6 | middle | 17.33±6.98 | 12.86±1.35 | 0.79 | 1425 | |
| 7 | Pm: Qv: Cf (7:2:1) | S | 10 | down | 15.56±4.59 | 12.66±1.23 | 0.80 | 1580 | |
| 8 | Pm: Qv: Cf (7:2:1) | E | 6 | down | 14.00±4.12 | 11.49±1.22 | 0.82 | 1635 | |
| LT | 9 | Pm: Qv: Cf (7:2:1) | NE | 9.5 | down | 14.67±4.37 | 11.94±1.23 | 0.72 | 1055 |
| 10 | Pm: Qv: Cf: Ic (6:2:1:1) | SE | 4.5 | middle | 14.06±4.83 | 11.82±1.19 | 0.75 | 1266 | |
| 11 | Pm: Qv: Cf (6:2:2) | S | 4.5 | down | 16.67±7.59 | 13.23±1.26 | 0.75 | 1302 | |
| 12 | Pm: Qv: Cf (7:2:1) | SE | 5 | down | 15.45±6.05 | 12.17±1.27 | 0.65 | 972 | |
| 13 | Pm: Qv: (7:3) | S | 4.5 | middle | 14.91±4.43 | 11.93±1.25 | 0.75 | 1569 | |
| 14 | Pm: Qv (7:3) | SW | 6.5 | middle | 14.41±4.97 | 12.02±1.20 | 0.79 | 1302 | |
| 15 | Pm: Qv: Cf (8:1:1) | E | 15.5 | down | 15.55±4.52 | 12.08±1.29 | 0.77 | 1149 | |
| MT | 16 | Pm: Qv (8:2) | N | 15.5 | down | 17.45±4.29 | 13.22±1.32 | 0.71 | 935 |
| 17 | Pm: Qv: Cf (6:3:1) | N | 11 | down | 14.26±4.16 | 11.42±1.25 | 0.80 | 1181 | |
| 18 | Pm: Qv (7:3) | N | 15.5 | down | 14.92±4.81 | 11.84±1.26 | 0.70 | 956 | |
| 19 | Pm: Qv (8:2) | E | 9.5 | down | 14.70±4.54 | 11.77±1.25 | 0.60 | 680 | |
| 20 | Pm: Qv (9:1) | S | 5 | up | 17.57±5.56 | 13.11±1.34 | 0.71 | 971 | |
| 21 | Pm: Qv: Cf (6:3:1) | W | 10 | down | 15.92±7.12 | 12.44±1.28 | 0.68 | 818 | |
| 22 | Pm: Qv (9:1) | E | 6.5 | up | 15.71±3.39 | 12.60±1.25 | 0.60 | 649 | |
| 23 | Pm: Qv (8:2) | S | 11 | middle | 17.72±6.36 | 13.32±1.33 | 0.75 | 894 | |
| 24 | Pm: Qv: Cf (7:2:1) | W | 9.5 | up | 15.43±4.78 | 11.96±1.29 | 0.65 | 726 | |
| 25 | Pm: Qv (8:2) | SE | 8.5 | up | 18.09±7.64 | 13.11±1.38 | 0.65 | 793 | |
| CK | 26 | Pm: Qv: Cf (6:3:1) | E | 10 | middle | 12.65±6.10 | 11.307±1.12 | 0.92 | 2121 |
| 27 | Pm: Qv: Cf: Pc (2:6:1:1) | E | 12.5 | middle | 11.31±4.45 | 12.64±1.06 | 0.90 | 1661 | |
| 28 | Pm: Qv: Cf (7:2:1) | E | 13.5 | down | 12.84±4.48 | 11.27±1.14 | 0.90 | 2052 |
Note: WT: weak thinning; LT: light thinning; MT: moderate thinning; CK: control; and the same applies to the other figures and tables. Pm: Pinus massoniana; Qv: Quercus variabilis; Cf: Cupressus funebris; Ic: Ilex chinensis; Pc: Pistacia chinensis. |
Table 2 Regenerated species associated with different thinning intensities |
| Family | Genus | Species | CK | WT | LT | MT |
|---|---|---|---|---|---|---|
| Cupressaceae | Cupressus | C. funebris | √ | √ | √ | √ |
| Euphorbiaceae | Vernicia | V. fordii | √ | √ | ||
| Euphorbiaceae | Triadica | T. sebifera | √ | √ | √ | |
| Euphorbiaceae | Mallotus | M. apelta | √ | √ | √ | √ |
| Euphorbiaceae | Glochidion | G. puberum | √ | √ | ||
| Aquifoliaceae | Ilex | I. cornuta | √ | |||
| Aquifoliaceae | Ilex | I. chinensis | √ | √ | √ | √ |
| Fabaceae | Dalbergia | D. hupeana | √ | √ | √ | |
| Fabaceae | Albizia | A. kalkora | √ | √ | ||
| Ericaceae | Rhododendron | R. simsii | √ | |||
| Eucommiaceae | Eucommia | E. ulmoides | √ | |||
| Pittosporaceae | Pittosporum | P. truncatum | √ | √ | √ | √ |
| Juglandaceae | Platycarya | P.strobilacea | √ | |||
| Fagaceae | Quercus | Q. variabilis | √ | √ | √ | √ |
| Meliaceae | Melia | M. azedarach | √ | |||
| Verbenaceae | Vitex | V. negundo | √ | √ | √ | |
| Oleaceae | Ligustrum | L. lucidum | √ | |||
| Anacardiaceae | Cotinus | C. coggygria | √ | |||
| Anacardiaceae | Rhus | R. chinensis | √ | √ | √ | √ |
| Anacardiaceae | Pistacia | P. chinensis | √ | √ | √ | |
| Rosaceae | Dichotomanthes | D. tristaniicarpa | √ | √ | √ | |
| Rosaceae | Prunus | P. salicina | √ | |||
| Moraceae | Broussonetia | B. papyrifera | √ | √ | √ | |
| Symplocaceae | Symplocos | S. sumuntia | √ | √ | √ | √ |
| Staphyleaceae | Euscaphis | E. japonica | √ | |||
| Ebenaceae | Diospyros | D. kaki | √ | √ | √ | |
| Ebenaceae | Diospyros | D. lotus | √ | √ | ||
| Rhamnaceae | Ziziphus | Z. jujuba | √ | |||
| Pinaceae | Pinus | P. massoniana | √ | √ | √ | |
| Ulmaceae | Celtis | C. sinensis | √ | √ | √ | |
| Rutaceae | Zanthoxylum | Z. avicennae | √ | |||
| Lauraceae | Lindera | L. glauca | √ | √ | √ | √ |
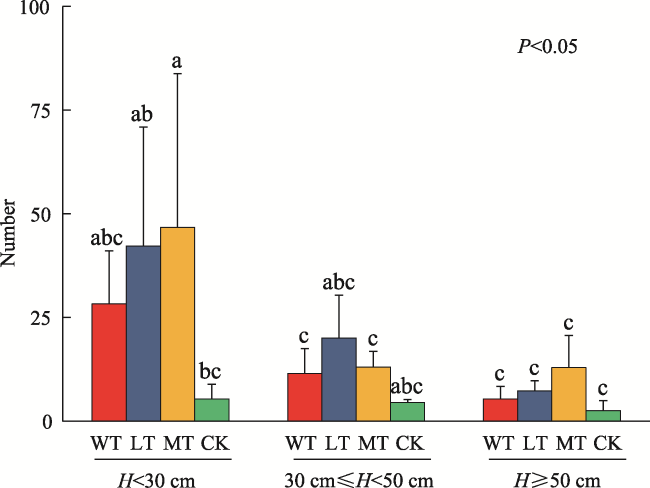
Fig. 1 Height distribution of regenerated trees in different thinning intensities |
Table 3 Natural regeneration seedling heights and regeneration densities at each thinning intensity |
| Thinning intensity | Different height class (H) regeneration densities (plants ha-1) | |||
|---|---|---|---|---|
| H<30 cm | 30 cm≤H<50 cm | H≥50 cm | Total | |
| CK | 2133 | 2000 | 533.2 | 4667 |
| WT | 11300 | 4600 | 2150 | 18050 |
| LT | 16857 | 8000 | 2914 | 27771 |
| MT | 18667 | 5200 | 5156 | 29022 |
Table 4 Richness and diversity of regenerated species in the undergrowth |
| Thinning intensity | S | D | H′ | Jsw | Ea |
|---|---|---|---|---|---|
| CK | 5.67±0.58b | 0.75±0.03a | 1.52±0.18a | 0.92±0.03a | 0.87±0.06a |
| WT | 7.38±2.20ab | 0.70±0.14a | 1.49±0.39a | 0.77±0.13b | 0.74±0.12ab |
| LT | 9.57±4.08a | 0.73±0.10a | 1.63±0.38a | 0.75±0.09b | 0.70±0.08b |
| MT | 9.55±2.55a | 0.70±0.11a | 1.54±0.24a | 0.73±0.13b | 0.68±0.15b |
Table 5 Multiple comparisons of intrinsic diversities of regenerated tree species |
| Thinning intensity | Species count | Intrinsic diversity higher than the stand | Intrinsic diversity lower than the stand |
|---|---|---|---|
| CK | 10 | CK≤WT, LT, MT | ≤CK |
| WT | 18 | WT≤LT, MT | CK≤WT |
| LT | 24 | LT≤MT | CK, WT≤LT |
| MT | 27 | MT≤ | CK, WT, LT≤MT |
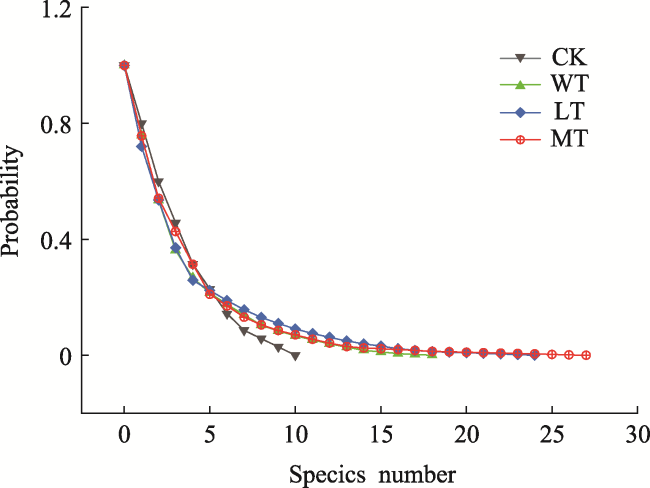
Fig. 2 The curves of right-tailed sums of diversities of regenerated tree species for the different thinning intensity levels |
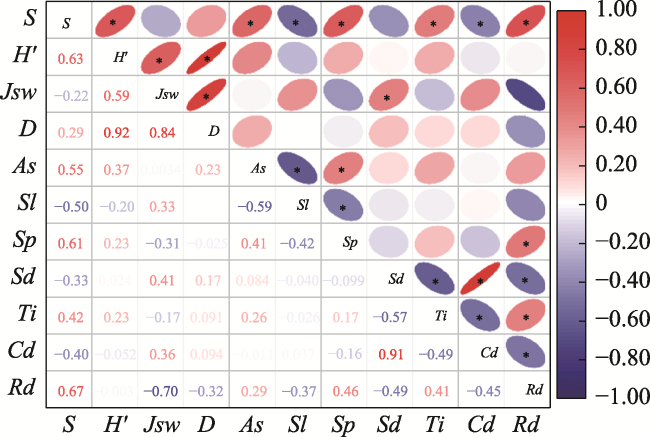
Fig. 3 Effects of stand factors and environmental factors on regenerationNote: P<0.05; S: Richness; H′: Shannon-Weiner index; Jsw: Pielou evenness; D: Simpson index; As: Aspect; Sl: slope; Sp: slope position; Sd: stand density; Ti: thinning intensity; Cd: canopy density; Rd: regeneration density. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}