
Journal of Resources and Ecology >
The Effects of Plateau Pika (Ochotona curzoniae) Presence and Population Control on the Structure of an Alpine Grassland Bird Community
|
Joseph P. LAMBERT, E-mail: joelangbo@yahoo.com |
Received date: 2021-05-20
Accepted date: 2022-03-20
Online published: 2023-01-31
Supported by
The National Natural Science Foundation of China(31470567)
China’s Qinghai-Tibetan Plateau (QTP) is an important area for bird conservation, with many endemic and Threatened species. Colonial burrowing mammals play an important role in structuring bird communities in arid grasslands around the world. On the QTP, the plateau pika Ocho tona curzoniae builds colonies which provide a dense source of resources for many bird species. However, pikas are regarded as a pest by local pastoralists, and they are the target of a population reduction campaign which could have a significant impact on the bird communities. We surveyed bird communities at Gansu Yanchiwan National Nature Reserve to investigate the differences in community structure between sites with pika colonies (on colony) and sites without them (off colony), and between pika colonies which had been poisoned and those which had not. Using non-metric multidimensional scaling (NMDS) combined with permutational multivariate analysis of variance (PERMANOVA) and Wilcoxon rank-sum tests, we found that there was no significant difference in bird community composition or abundance between the poisoned and untreated colonies. However, there was a very large and statistically significant difference in bird community structures between on- and off-colony sites. Only horned lark Eremophila alpestris was consistently observed at sites without pika colonies, while ten bird species were observed on colonies. Six species were significantly more abundant on colony than off. While we could not claim that the poisoning campaign at Yanchiwan is altering bird communities, the presence of pika colonies seems to be an indispensable resource for the resident birds.
Key words: birds; community ecology; ecosystem engineer; plateau pika; Qinghai-Tibetan Plateau
Joseph P. LAMBERT , Johanna V. HARTMANN , SHI Kun , Philip RIORDAN . The Effects of Plateau Pika (Ochotona curzoniae) Presence and Population Control on the Structure of an Alpine Grassland Bird Community[J]. Journal of Resources and Ecology, 2023 , 14(1) : 46 -56 . DOI: 10.5814/j.issn.1674-764x.2023.01.005
Fig. 1 Location of Gansu Yanchiwan NNR within China’s Gansu Province |
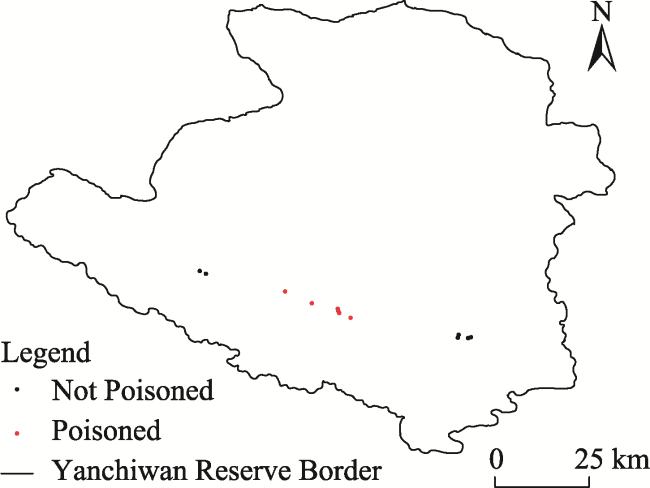
Fig. 2 Map of poisoned and untreated (i.e., not poisoned) survey sites in Yanchiwan NNR for 2017 |
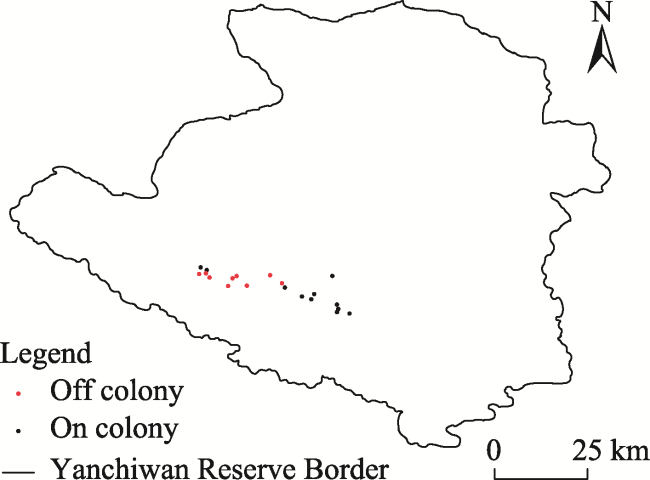
Fig. 3 Map of survey sites on and off pika colonies surveyed in 2018 in Yanchiwan NNR |
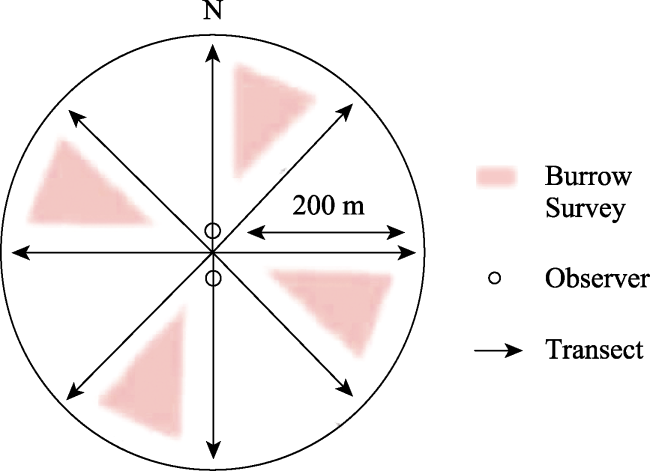
Fig. 4 Schematic diagram of the site and transects |
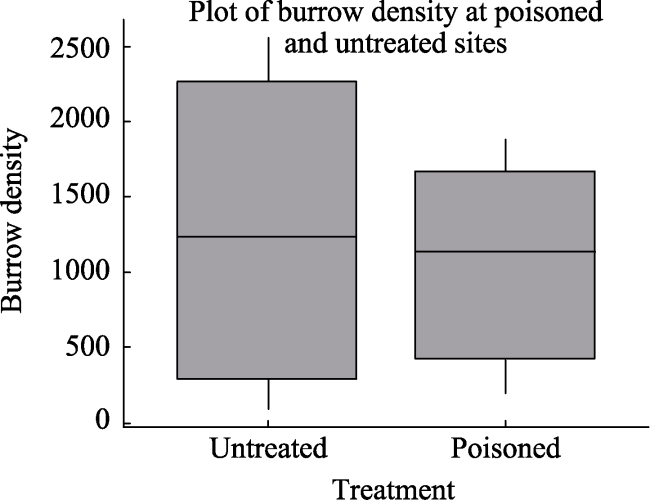
Fig. 5 Box plot showing burrow densities at poisoned and untreated sites |
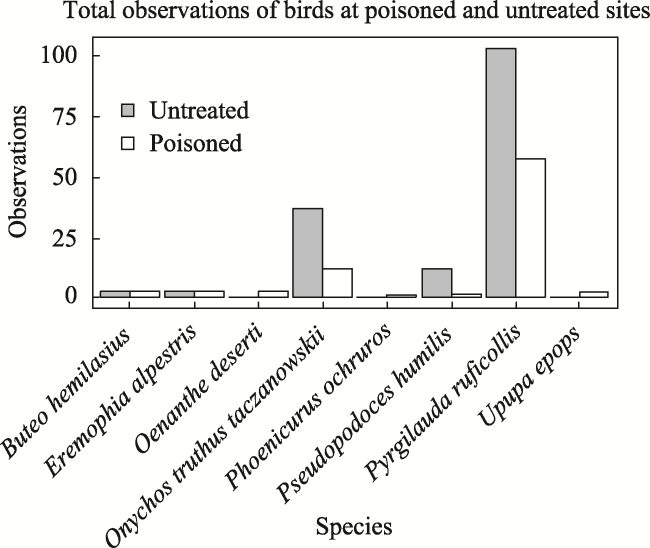
Fig. 6 Bar chart showing total observations of each bird species at poisoned and untreated sites in 2017Note: None of the differences were significant according to Wilcoxon rank sum tests (P >0.05). |
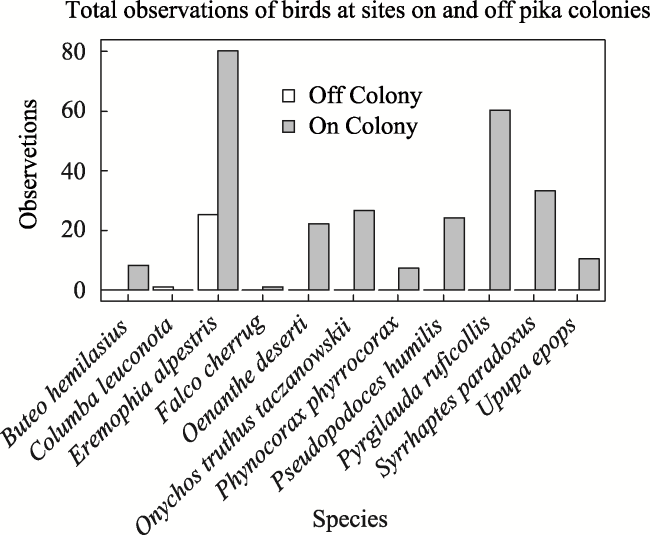
Fig. 7 Graph of total observations of bird species at sites off and on pika colonies in 2018Note: Differences were significant for P. humilis (W = 72, P = 0.02), U. epops (W = 72, P = 0.02), S. paradoxus, (W = 72, P = 0.02), P. ruficollis (W = 90, P = <0.001), O. taczanowskii (W = 72, P = 0.02) and B. hemilasius (W = 76.5, P = 0.01). |
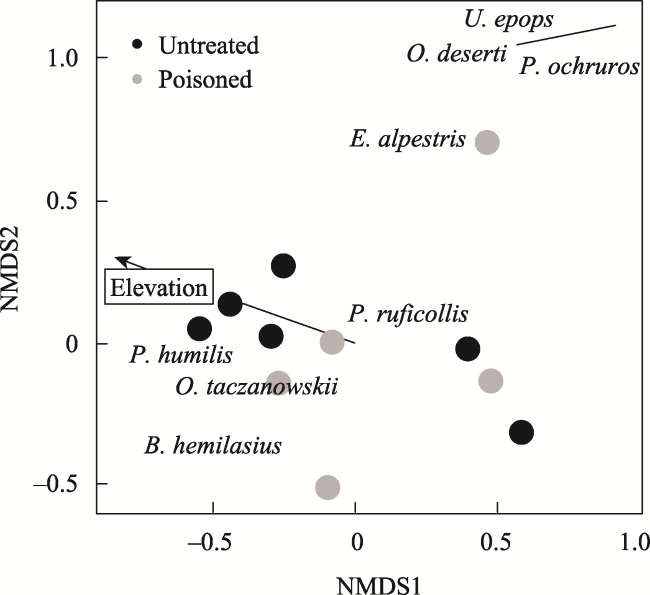
Fig. 8 NMDS of bird communities at poisoned and untreated sites during 2017Note: Envfit vector displayed for elevation (R2 = 0.7, P = 0.01). |
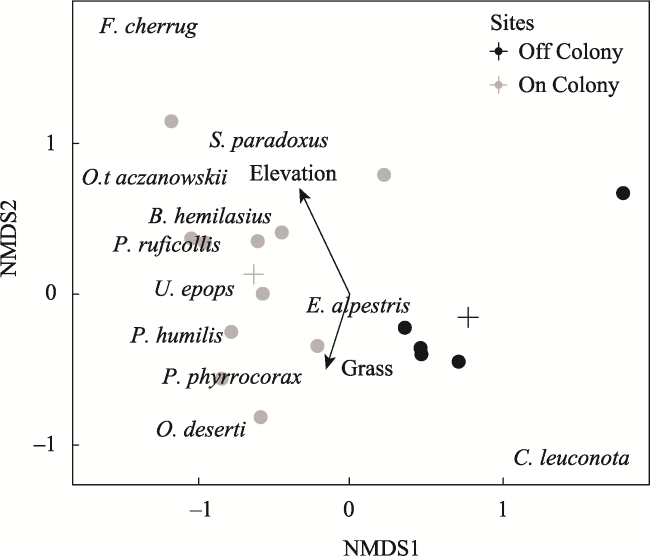
Fig. 9 NMDS of bird communities at sites on and off pika colonies in 2018Note: Crosses represent significantly different centroid locations for categorical variables according to PERMANOVA (F = 7.32, P < 0.001). Envfit vectors are plotted for variables where the correlation was significantly different from zero (i.e. elevation, R2 = 0.5, P = 0.001) and grass cover (R2 = 0.2, P = 0.05). |
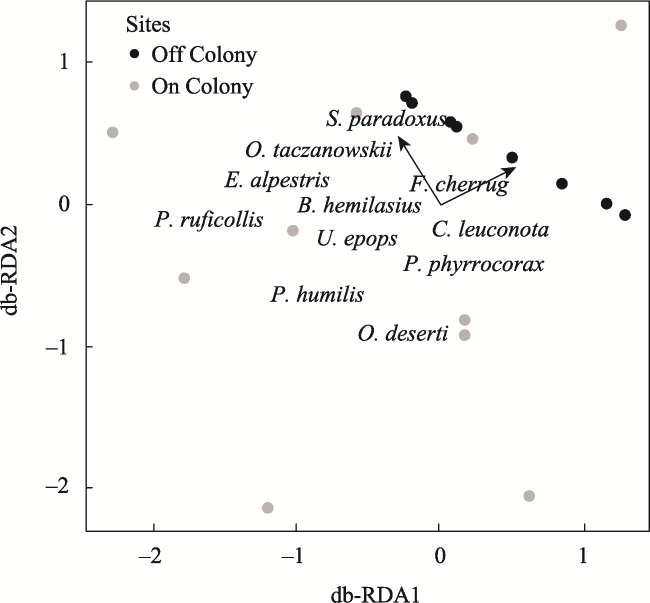
Fig. 10 Plot of the partial distance-based redundancy analysis (partial db-RDA) of bird communities at on-colony and off-colony sites in 2018Note: Statistically significant vectors plotted for elevation (P = 0.047) and plant richness (P = 0.019). |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
Badingqiuying. 2016. Pikas, grasslands, and pastoralists: Understanding the roles of plateau pikas in a coupled social-ecological system. Diss., Phoenix, USA: Arizona State University.
|
| [5] |
Badingqiuying,
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
Bird Life International. 2009. Directory of important bird areas in China (Mainland): Key sites for conservation. Cambridge, UK: Bird Life International.
|
| [10] |
Bird Life International. 2017. The IUCN red list of threatened species 2017. DOI: 10.2305/IUCN.UK.2017-1.RLTS.T22696495A110525916.en.
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
Van Nimwegen R E,
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}