
Journal of Resources and Ecology >
Effects of Prosopis juliflora Invasion on Native Species Diversity and Woody Species Regenerations in Rangelands of Afar National Regional State, Northeast Ethiopia
Received date: 2021-07-25
Accepted date: 2022-06-29
Online published: 2023-01-31
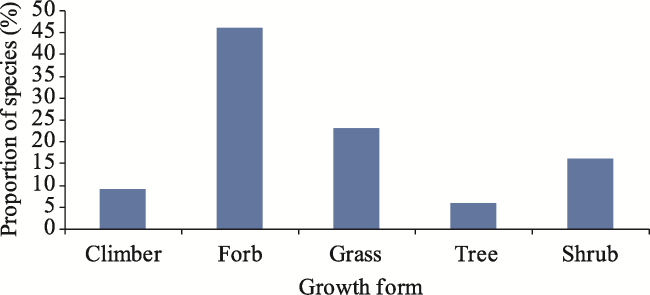
Investigation of the invasion of Prosopis juliflora and its effects on indigenous plant species are important for the control of the species. The study aimed to assess: (1) the effects of Prosopis juliflora invasion on the diversity of plant species in Awash Fentale and Amibara Woredas; (2) the effects of Prosopis juliflora invasion on the regeneration potential of native woody species. Sample collection was performed in habitats of Prosopis juliflora thicket, Prosopis juliflora mixed with native species stands, non-invaded woodlands, and open grazing lands. The vegetation was stratified into invasion levels of Prosopis juliflora and then a random sampling technique for data collection. Among species of plants, the highest proportion of species, 75 (47.8%), was recorded under non-invaded woodlands, but the lowest proportion of species, 22 (14%), was recorded under open grazing lands. The invasion of Prosopis juliflora reduced the Shannon diversity index. The mean values of the Shannon diversity index and species richness under Prosopis juliflora mixed with native species (H´=2.22, R=14) and non-invaded woodlands (H´=2.23, R=13) were significantly higher than Prosopis juliflora thicket (H´=1.96, R=12) and open grazing lands (H´=1.84, R=10). The highest total density (358 stems ha‒1) of seedlings was recorded under Prosopis juliflora mixed with native species. But, the lowest total density (153 stems ha‒1) of seedlings was recorded under Prosopis juliflora thickets. Moreover, 102 trees ha‒1 native woody species were recorded under Prosopis juliflora thicket, but 1252 trees ha‒1 native species were recorded under non-invaded woodlands. If the invasion of Prosopis juliflora and its effects on native species diversity continue coupled with a drier climate, plant diversity of the Afar flora region will be highly affected and its ecosystem services will be under question. Thus, the participation of all stakeholders and multidisciplinary research approaches should be designed for the management of the species and rehabilitation of the rangelands in the region.
Wakshum SHIFERAW , Sebsebe DEMISSEW , Tamrat BEKELE , Ermias AYNEKULU . Effects of Prosopis juliflora Invasion on Native Species Diversity and Woody Species Regenerations in Rangelands of Afar National Regional State, Northeast Ethiopia[J]. Journal of Resources and Ecology, 2023 , 14(1) : 35 -45 . DOI: 10.5814/j.issn.1674-764x.2023.01.004
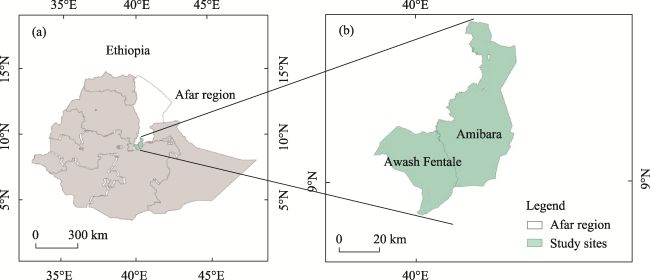
Fig. 1 Location of the study area (Afar region) in Ethiopia (a) and location of Amibara Woreda and Awash Fentale Woreda in Afar region (b) |
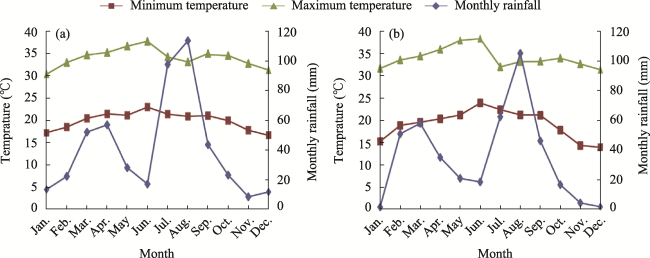
Fig. 2 Thirty-one-year (1986-2017) climate diagram for Awash Fentale Woreda (a) and fifteen-year (1986-2001) climate diagram for Amibara Woreda (b)Note: Data source: Shiferaw et al. (2020). |
Fig. 3 Plates showing P. juliflora invasion levels or habitats in Afar Floristic RegionNote: Plate (a) indicates an area invaded purely by P. juliflora and plate (b) P. juliflora mixed with native species, however, plate (c) is non-invaded woodlands without P. juliflora invasion and plate (d) is only grassland area not invaded by P. juliflora or encroached by other native woody species. |
Fig. 4 Proportion (%) of plant species in the study woredas |
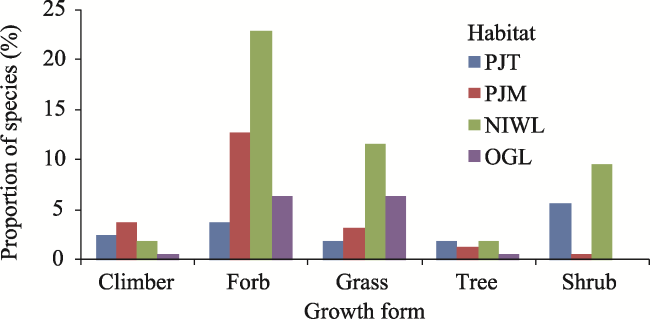
Fig. 5 Proportion (%) of plant species in each habitatNote: PJT is Prosopis juliflora thicket; PJM is Prosopis juliflora mixed with native species; NIWL is non-invaded woodland; OGL is open grazing land. P. juliflora invasions levels didn’t show significant variations in growth forms (P > 0.05). |
Table 1 GLM showing effects location and P. juliflora invasion levels on species diversity in Amibara and Awash Fentale Woredas, Ethiopia |
| Explanatory variable ×Response variable | Source | df | Sum of squares | Mean square | F-value | P-value (Pr >F) |
|---|---|---|---|---|---|---|
| Model | 3 | 1.86 | 0.62 | 3.74 | 0.016 | |
| Habitat ×H° | Error | 60 | 9.92 | 0.17 | - | - |
| Corrected total | 63 | 11.77 | - | - | - | |
| Model | 3 | 191.42 | 63.81 | 3.52 | 0.020 | |
| Habitat × R | Error | 60 | 1086.81 | 18.11 | - | - |
| Corrected total | 63 | 1278.23 | - | - | - | |
| Model | 3 | 0.01997 | 0.0067 | 2.41 | 0.076 | |
| Habitat × E° | Error | 60 | 0.166 | 0.0028 | - | - |
| Corrected total | 63 | 0.186 | - | - | - | |
| Model | 1 | 0.068 | 0.068 | 0.26 | 0.61 | |
| District × H° | Error | 62 | 15.94 | 0.26 | ||
| Corrected total | 63 | 16.006 | ||||
| Model | 1 | 0.0 | 0.0 | 0.0 | 1.0 | |
| District × R | Error | 62 | 4716.0 | 76.06 | ||
| Corrected total | 63 | 4716.0 | ||||
| Model | 1 | 0.005 | 0.0049 | 0.33 | 0.57 | |
| District × E° | Error | 62 | 0.948 | 0.0153 | ||
| Corrected total | 63 | 0.9527 |
Note: Significant at α = 0.05, H° is Shannon diversity, R is species richness, E° is Shannon evenness, Pr > F is the P-value associated with the F statistic of a given effect and test statistic. The null hypothesis that a given predictor has no effect on either of the outcomes is evaluated with regard to this P-value. |
Table 2 Mean values of vegetation patterns for P. juliflora invasion levels in Awash Fentale and Amibara Woredas |
| Vegetation patterns | Prosopis invasion levels | |||
|---|---|---|---|---|
| PJT (N=16) | PJM (N=16) | NIWL (N=16) | OGL (N=16) | |
| Shannon diversity (H°) | 1.96ab | 2.22a | 2.23a | 1.84b |
| Richness (R) | 11.50ab | 13.94a | 13.44a | 9.56b |
| Shannon evenness (E´) | 0.83a | 0.86a | 0.87a | 0.83a |
| Vegetation patterns | District | |||
| Awash Fentale (N= 32) | Amibara (N=32) | |||
| Shannon diversity (H°) | 1.96a | 1.90a | ||
| Richness (R) | 58a | 58a | ||
| Shannon evenness (E°) | 0.49a | 0.47a |
Note: PJT is P. juliflora thicket; PJM is mixed (P. juliflora + native woody species); NIWL is non-invaded woodland; OGL is open grazing land; N= Number of individuals per 400 m2. Values with similar letters are insignificant (P >0.05), but different letters are significantly different (P <0.05). |
Table 3 General linear model showing the effects of P. juliflora invasion levels on regeneration status of woody species in Awash Fentale and Amibara Woredas |
| Response variables | Source | df | Sum of squares | Mean square | F-value | P-value (Pr> F) |
|---|---|---|---|---|---|---|
| Model | 2 | 36907.7 | 18454 | 0.94 | 0.41 | |
| Habitat × Trees | Error | 21 | 410318.9 | 19539 | - | - |
| Corrected total | 23 | 447226.7 | ||||
| Model | 2 | 32.66 | 16.33 | 0.71 | 0.49 | |
| Habitat × Saplings | Error | 1592 | 36640.01 | 23.01 | - | - |
| Corrected total | 1594 | 36672.67 | - | - | - | |
| Model | 2 | 69.28 | 34.64 | 1.06 | 0.35 | |
| Habitat × Seedlings | Error | 1592 | 51796.23 | 32.53 | - | - |
| Corrected total | 1594 | 51865.51 | - | - | - |
Note: α = 0.05, Pr > F is the P-value associated with the F statistic of a given effect and test statistic. The null hypothesis that a given predictor has no effect on either of the outcomes is evaluated with regard to this P-value. |
Table 4 Mean values of the regeneration status for P. juliflora invaded and non-invaded habitats at Amibara and Awash Fentale districts, Ethiopia (Unit: stems ha-1) |
| Growth Stages | Habitat | ||
|---|---|---|---|
| PJT | PJM | NIWLs | |
| Trees | 102a | 585b | 1252b |
| Saplings | 151a | 334b | 324c |
| Seedlings | 153a | 358b | 242c |
Note: PJT is P. juliflora thicket; PJM is P. juliflora mixed with native species; NIWLs is non-invaded woodlands. |
Table 6 Appendix 1 Plant species list in Awash Fentale and Amibara Woredas |
| Scientific name | Family | Life form | Vernacular name* | SCN |
|---|---|---|---|---|
| Abutilon anglosomaliae Cufod. | Malvaceae | Forb | Hambukto | WSC143 |
| Abutilon figarianum Webb | Malvaceae | Forb | Hedayito | WSC154 |
| Abutilon fruticosum Guill. & Perr. | Malvaceae | Shrub | Hambukto | WSC005 |
| Abutilon ramosum Guill. & Perr. | Malvaceae | Forb | Hambukto | WSC144 |
| Acacia mellifera (Vahl) Benth. | Fabaceae | Shrub | Makharto | WSC007 |
| Acacia nilotica (L.) Willd. ex. Del. | Fabaceae | Tree | Kasalto | WSC145 |
| Acacia oerfota (Forssk) Schweinf. | Fabaceae | Tree | Gerento | WSC115 |
| Acacia senegal (L.) Wild. | Fabaceae | Tree | Adado | WSC104 |
| Acacia tortilis (Frossk.) Hayne | Fabaceae | Tree | E’ebto | WSC003 |
| Acalypha acrogyna Pax | Euphorbiaceae | Forb | Berbere | WSC063 |
| Achyranthes aspera L. var. pubeseens Moq. | Amaranthaceae | Forb | Murit tabiri | WSC125 |
| Aerva javanica (Burm.f.) Schultes | Amaranthaceae | Shrub | Olyato | WSC018 |
| Alternanthera pungens Kunth. | Amaranthaceae | Forb | Ferengisisib | WSC040 |
| Amaranthus dubius Thell. | Amaranthaceae | Forb | Bonket/Bunkete | WSC146 |
| Amaranthus thunbergii Moq. | Amaranthaceae | Forb | Aburi | WSC107 |
| Asparagus africanus Lam. | Asparagaceae | Forb | ‒ | WSC147 |
| Balanites aegyptiaca (L.) Del. | Balanitaceae | Tree | Udda | WSC044 |
| Barleria acanthoides Vahl | Acanthaceae | Forb | Ganzalto | WSC004 |
| Berchemia discolor (Klotzsch) Hemsl. | Rhamnaceae | Tree | Jajaba'ito | WSC122 |
| Blepharis maderaspatensis (L.) Roth | Acanthaceae | Forb | Yamarukta | WSC118 |
| Boerhavia coccinea Mill. | Nyctaginaceae | Forb | Asara | WSC167 |
| Boerhavia repens L. | Nyctaginaceae | Forb | Asara | WSC119 |
| Brachiaria ramosa (L.) Stapf | Poaceae | Herb (A) | Gewita | WSC075 |
| Cadaba glandulosa Forssk. | Capparidaceae | Shrub | Ududoyta | WSC139 |
| Cadaba longifolia (R. Br.) DC. | Capparidaceae | Shrub | Dunbiya | WSC110 |
| Cadaba rotundifolia Forssk. | Capparidaceae | Tree | Haragali | WSC184 |
| Calotropis procera (Ait.) Ait. f. | Asclepiadaceae | Forb | Gelato/garbaadiyita | WSC059 |
| Capparis tomentosa Lam. | Capparidaceae | Climber | Dangayito | WSC169 |
| Ceinfugosia somaliana Fryx | Malvaceae | Shrub | Hameresa | WSC117 |
| Cenchrus ciliaris L. | Poaceae | Herb (P) | Serdoyita | WSC024 |
| Centrostachys aquatica (R.Br.) Wall. ex Moq. | Amaranthaceae | Forb | Bete | WSC038 |
| Chenopodium album L. | Chenopodiaceae | Forb | Riba | WSC073 |
| Chrysopogon plumulosus Hochst. | Poaceae | Herb (P) | Durfu | WSC079 |
| Cissus quadrangularis L. | Vitaceae | Climber | Ala’e | WSC149 |
| Cissus rotundifolia (Forssk.) Vahl | Vitaceae | Climber | Al’ie | WSC106 |
| Clematis hirsuta Perr. & Guill. | Ranunculaceae | Climber | Adayito | WSC056 |
| Clematis longicauda Steud. ex A. Rich. | Ranunculaceae | Climber | Matamato | WSC172 |
| Cocculus pendulus (J.R. & G. Forst) | Menispermaceae | Climber | Hayikto | WSC131 |
| Commelina diffusa Burm.f | Commelinaceae | Forb | Mutuki | WSC070 |
| Commicarpus plumbagineus (Cav.) Standl. | Nyctaginaceae | Climber | Yejib chama | WSC162 |
| Commiphora coronillifolia Chiov. | Burseraceae | Shrub | File neme’a | WSC033 |
| Cordia sinensis Lam. | Boraginaceae | Shrub | Maderto | WSC002 |
| Cryptostegia grandiflora Roxb. Ex R. Br. | Asclepiadaceae | Climber | Halimaro | WSC046 |
| Cucumis dipsaceus Ehrnb. ex spach. | Cucurbitaceae | Climber | Seroyita | WSC051 |
| Cucumis prophetarum L. | Cucurbitaceae | Forb | Garaun | WSC089 |
| Cyathula cylindrica Moq. | Amaranthaceae | Forb | Kandadaf | WSC077 |
| Cycnium erectum Rendle | Sclrofloriaceae | Forb | Andoliva | WSC151 |
| Cymbopogon commutatus (Steud.) Stapf | Poaceae | Herb (P) | Haragali | WSC083 |
| Cymbopogon pospischilii (K. Schum.) C.E. Hubb. | Poaceae | Herb | ‒ | WSC170 |
| Cynodon dactylon (L.) Pers. | Poaceae | Herb (P) | Rareta | WSC041 |
| Cynodon nlemfuensis Vanderyst | Poaceae | Herb (P) | Rareta | WSC037 |
| Dactyloctenium scindicum Boiss. | Poaceae | Herb (P) | Afara mole | WSC026 |
| Delonix elata L. Gamble | Fabaceae | Tree | Amayito | WSC134 |
| Diceratella elliptica Guill. & Perr. | Brassicaceae | Herb | Moroe’i | WSC080 |
| Diceratella incana Balf.f. | Brassicaceae | Forb | Atikhara | WSC103 |
| Digitaria velutina (Forssk.) P. Beauv. | Poaceae | Herb (A) | Santkefa | WSC097 |
| Dobera glabra (Forssk.) Poir. | Salvadoraceae | Shrub | Gersayiato | WSC154 |
| Dregea rubicunda K. Schum. | Asclepiadaceae | Forb | Golfaha | WSC064 |
| Ecbolium viride (Forssk.) Alston | Acanthaceae | Forb | Kinoyita | WSC120 |
| Echinochloa colona (L.) Link | Poaceae | Herb (A) | ‒ | WSC163 |
| Eleusine africana Kenn.‒O’Byrne | Poaceae | Herb (A) | ‒ | WSC110 |
| Eragrostis aethiopica Chiov. | Poaceae | Herb (A) | Ayti‒adoyta | WSC074 |
| Eragrostis biflora Hack. | Poaceae | Herb (A) | ‒ | WSC155 |
| Eragrostis cilianensis (All.) Vign. ex Janchen | Poaceae | Herb | ‒ | WSC118 |
| Eragrostis cylindriflora Hochst. | Poaceae | Herb (A) | Dankit/feresgera | WSC156 |
| Eragrostis papposa (Roem. & Schult.) Steud. | Poaceae | Herb (P) | Bekelayso | WSC078 |
| Eriochloa fatmensis (Hochst. & Steud.) Clayton | Poaceae | Herb (A) | Bohale | WSC019 |
| Erucastrum arabicum Fisch. & Mey. | Brassicaceae | Forb | Ferate | WSC026 |
| Eurphorbia longituberculosa Boiss. | Euphorbiaceae | Forb | Haliforesi | WSC034 |
| Fagonia schweinfurthii (Hadidi) Hadidi. | Zagophyllaceae | Shrub | Adihara | WSC129 |
| Fuirena leptostachya Oliv. | Cyperaceae | Herb | Ka’ato | WSC158 |
| Glossonema revoilii Franch. | Asclepiadaceae | Forb | Sanga hayu | WSC081 |
| Grewia flavescens Juss. | Tiliaceae | Shrub | Menangure/Garuwayito | WSC035 |
| Grewia schweinfurthii Burret | Tiliaceae | Shrub | Manangurita/AdePJTo | WSC127 |
| Grewia tenax (Forssk.) Fiori | Tiliaceae | Forb | Hedayito | WSC032 |
| Grewia villosa Willd. | Tiliaceae | Shrub | Garawayito/legida | WSC035 |
| Heliotropium longiflorum (A.DC. in DC.) Jaub. & Spach | Boraginaceae | Forb | Kinbira hara | WSC092 |
| Hermannia paniculata Franch. | Sterculiaceae | Shrub | ‒ | WSC043 |
| Hibiscus calyphyllus Cavan. | Malvaceae | Shrub | Hambukito | WSC091 |
| Hibiscus crassinervius Hochst. ex A. Rich. | Malvaceae | Forb | Akulito | WSC106 |
| Hibiscus dongolensis Del. | Malvaceae | Forb | Walayneba | WSC028 |
| Hibiscus micranthus L. f. | Malvaceae | Forb | Okelehina | WSC164 |
| Huernia somalica N.E. Br. | Asclepiadaceae | Forb | ‒ | WSC200 |
| Hyparrhenia rufa (Nees) Stapf | Poaceae | Herb (P) | Isesu | WSC163 |
| Hypertelis bowkeriana Sond. | Molluginaceae | Forb | Asara | WSC073 |
| Hypoestes forskaolii (Vahl) R. Br. | Acanthaceae | Forb | Harawayitu | WSC006 |
| Hypoestes triflora (Forssk.) Roem. & Schult. | Acanthaceae | Forb | Harawayitu | WSC180 |
| Hyptis pectinata (L.) Poit. | Lamiaceae | Forb | Amada | WSC165 |
| Hyptis spicigera Lam. | Lamiaceae | Forb | Amado | WSC054 |
| Indigofera coerulea Roxb. | Fabaceae | Forb | Dunwayito | WSC126 |
| Indigofera hochstetteri Bak. | Fabaceae | Forb | Aro | WSC060 |
| Ipomoea cairica (L.) Sweet | Convolvulaceae | Climber | ‒ | WSC166 |
| Jatropha ellenbeckii Pax | Euphorbiaceae | Forb | ‒ | WSC036 |
| Jatropha glauca Vahl | Euphorbiaceae | Shrub | Halfolisi | WSC022 |
| Kleinia odora (Forssk.) DC. | Astreraceae | Climber | Bisilto | WSC133 |
| Leptadenia hastata (Pers.) Decne. | Asclepiadaceae | Climber | Hayikto | WSC012 |
| Leucas inflata Benth | Lamiaceae | Forb | Bunketi | WSC025 |
| Leucas martinicensis (Jacq.) R. Br. | Lamiaceae | Forb | Kurufule | WSC160 |
| Linum strictum L. | Linaceae | Forb | ‒ | WSC090 |
| Linum volkensii Engl. | Linaceae | Forb | Susui | WSC140 |
| Lipocarpha hemisphaericus (Roth) Goetgh. | Cyperaceae | Herb | Gerandoyta/Gedoyta | WSC153 |
| Lipocarpha rehmannii (Ridl.) Goetgh. | Cyperaceae | Herb | Abuu | WSC111 |
| Lonchocarpus laxiflorus Guill. & Perr. | Fabaceae | Forb | Halemagira | WSC143 |
| Maerua angolensis DC. | Capparidaceae | Shrub | Dunbayito | WSC144 |
| Megalochlamys ogadenensis Vollesen | Acanthaceae | Shrub | Gelsanto | WSC116 |
| Megalochlamys violacea (Vahl) Vollesen | Acanthaceae | Forb | Uraurto | WSC052 |
| Otostegia fruticosa (Forssk.) Schweinf.ex Penzig | Acanthaceae | Forb | Atihara/michi | WSC169 |
| Monothecium glandulosum Hochst. | Acanthaceae | Forb | Kulumintili | WSC069 |
| Nicotiana glauca R. Grah. | Solanaceae | Forb | Adihara | WSC170 |
| Ocimum circinatum A.J Paton | Lamiaceae | Forb | Harawayitu | WSC106 |
| Ocimum forskolei Benth | Lamiaceae | Forb | Kayidiriya | WSC085 |
| Ocimum jamesii Sebald | Lamiaceae | Herb | ‒ | WSC174 |
| Ocimum americanum L. | Lamiaceae | Forb | Kaqtuhara | WSC016 |
| Orthosiphon pallidus Royle ex Benth. | Lamiaceae | Forb | Yengula hebaki | WSC067 |
| Otostegia fruticosa (Forssk.) Schweinf. ex Penzig | Lamiaceae | Shrub | ‒ | WSC183 |
| Otostegia modesta S. Moore | Lamiaceae | Forb | ‒ | WSC121 |
| Echinochloa colona (L.) Link | Poaceae | Herb (A) | Baruri | WSC034 |
| Panicum subalbidum Kunth | Poaceae | Herb (A) | Heniso | WSC124 |
| Parkinsonia scioana (Chiov). Brenan | Fabaceae | Shrub | Sosokite | WSC171 |
| Parthenium hysterophorus L. | Astreraceae | Forb | Hari | WSC001 |
| Paspalidium desertorum (A. Rich.) Stapf | Poaceae | Herb (P) | ‒ | WSC019 |
| Pavonia arabica Hochst. & Steud. ex Boiss. | Malvaceae | Forb | ‒ | WSC091 |
| Pergularia daemia (Forssk.) Chiov. | Asclepiadaceae | climber | Gime’ito | WSC076 |
| Peristrophe paniculata (Forssk.) Brummitt | Acanthaceae | Forb | Aburi | WSC136 |
| Pisonia aculeata L. | Nyctaginaceae | Climber | Dambikiso | WSC042 |
| Portulaca quadrifida L. | Portulacaceae | Forb | Hatihara | WSC175 |
| Prosopis juliflora (Sw.) DC. | Fabaceae | Shrub | Dargi‒haraa | WSC010 |
| Pupalia lappacea (L.) A. Juss. | Amaranthaceae | Forb | Sarotkefa | WSC024 |
| Salvadora persica L. | Salvadoraceae | Shrub | Adaito | WSC056 |
| Sansevieria ehrenbergii Schweinf. ex Baker. | Dracaenaceae | Forb | Ya’ato | WSC176 |
| Sansevieria forskaoliana (Schult. f.) Hepper & Wood | Dracaenaceae | Forb | Ya’a | WSC123 |
| Schoenoplectus maritimus (L.) Lye | Cyperaceae | Herb | Ka’ato | WSC053 |
| Seddera bagshawei Rendle. | Convolvulaceae | Forb | Riba | WSC023 |
| Seddera latifolia Hochst. & Steud. | Convolvulaceae | Forb | Adiriba | WSC114 |
| Senna italica Mill. | Fabaceae | Shrub | Oklehina | WSC027 |
| Senna obtusifolia (L.) Irwin & Barneby | Fabaceae | Shrub | Salilimeki | WSC109 |
| Setaria verticillata (L.) P. Beauv. | Poaceae | Herb (A) | Delayta | WSC177 |
| Solanum coagulans Forssk. | Solanaceae | Forb | Alulis | WSC178 |
| Solanum cordatum Forssk. | Solanaceae | Forb | Ubabulto | WSC050 |
| Solanum incanum L. | Solanaceae | Forb | Kurara’i | WSC066 |
| Solanum schimperianum Hochst. ex A. Rich. | Solanaceae | Forb | Bobao | WSC015 |
| Sporobolus agrostoides Chiov. | Poaceae | Herb (P) | ‒ | WSS171 |
| Sporobolus consimilis Fresen. | Poaceae | Herb (P) | Denekto | WSC180 |
| Sporobolus panicoides A.Rich. | Poaceae | Herb (A) | Gewita | WSC009 |
| Sporobolus pellucidus Hoehst. | Poaceae | Herb (P) | Sosokete | WSC181 |
| Sporobolus spicatus (Vahl) Kunth | Poaceae | Herb (P) | Edolatyansi | WSC039 |
| Tetrapogon cenchriformis (A.Rich.) Clayton | Poaceae | Herb (A) | Sabrisi | WSC045 |
| Thunbergia ruspolii Lindau | Acanthaceae | Forb | Harawayito | WSC086 |
| Tragus racemosus (L.) All. | Poaceae | Herb (A) | Bekelayso | WSC078 |
| Tribulus parvispinus Presl. | Zygophyllaceae | Forb | Bunket | WSC061 |
| Tribulus terrestris L. | Zygophyllaceae | Forb | Bunket | WSC182 |
| Verbesina encelioides (Cav.) A. Gray | Asteraceae | Forb | Surimia | WSC062 |
| Wissadula rostrata (Schumach. & Thonn.) Hook.f. | Malvaceae | Shrub | Delgida | WSC142 |
| Xanthium strumarium L. | Asteraceae | Forb | Bangi | WSC030 |
| Ziziphus spina‒christi (L.) Desf. var. mitissima Chiov. | Rhamnaceae | Tree | Kusirayito | WSC132 |
| Zygophyllum simplex L. | Zygophllaceae | Forb | Mutiki | WSC034 |
Note: Vernacular name* is plant name in Afar language. SCN is species collection name; A is annual grasses; P is perennial grasses. |
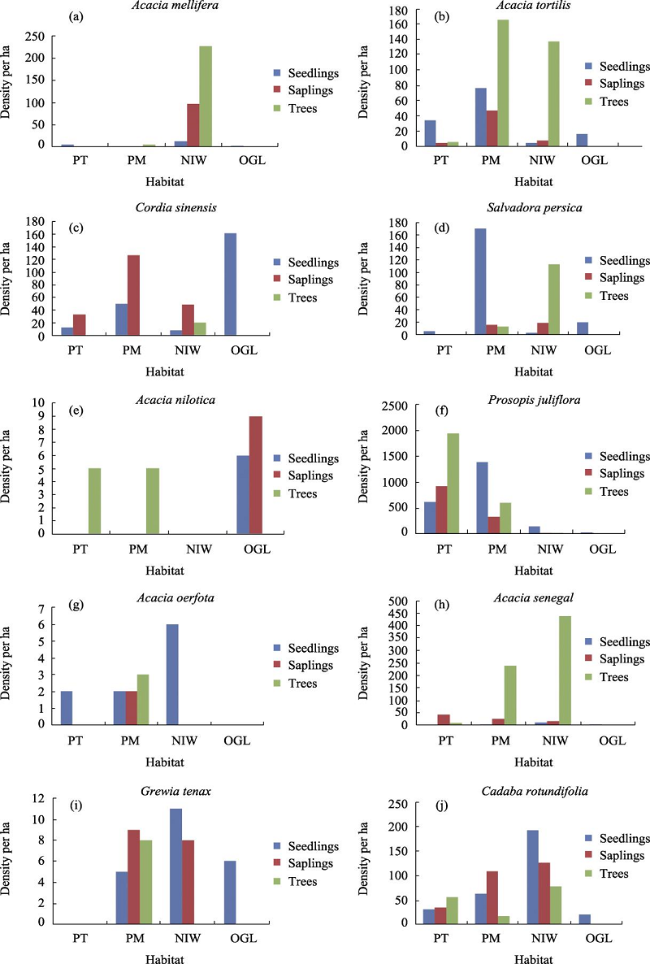
Fig 6 Appendix 2 a‒j. Size class structures of ten species population in each habitatNote: PT = P. juliflora thicket stands; PJM = P. juliflora with native species stands; NIWL = Non-invaded woodland; OGL = Open grazing land. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
Central Statistical Agency(CSA). 2013. Population projection of Ethiopia for all regions at district level, Addis Ababa, Ethiopia. www.sciepub.com.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
SAS. 2002. SAS/STAT user’s guide, version 9.0. Cary, USA: SAS Institute. sasdoc_91 › stat_ug_7313.
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}