
Journal of Resources and Ecology >
The Spatio-temporal Patterns of Macro Benthos Functional Groups and the Associated Factors Affecting Them after Wetland Restoration
|
WANG Maoqiu, E-mail: wangmaoqiu@yeah.net |
Received date: 2021-10-08
Accepted date: 2022-03-20
Online published: 2022-10-12
Supported by
The National Key Research and Development Program of China(2017YFC0506002)
The Foundation of Shanghai Marine Environmental Monitoring Center(MEMRT202003)
This study examines how the spatiotemporal patterns of functional groups of macro benthos responded to coastal wetland restoration projects. Compared with the traditional single-species approach, methods for identifying functional groups of macro benthos more comprehensively reflect the states of the wetland, i.e., spatiotemporal patterns and the related influencing factors. In this study, the macro benthos samples, soil samples, and plant samples were collected at each same sample site in April, July, and October of 2017 and January of 2018. After classification of the macro benthos functional groups according to some traits, the factors influencing the functional groups were examined by a linear stepwise regression. The results showed that all macro benthos were classified into 11 different functional groups based on their traits of forms of locomotion, feeding habits, and food gathering methods. The semi-mobile suspensivores (FDX), semi-mobile surface detritivores (SDX), and Jawed mobile carnivores (CMJ) were the main groups observed in a year. Regression analysis showed that particle size, moisture content, and plant height were important common factors influencing most groups. The main influencing factor of FDX was particle size (P=0.020). Moisture content (P=0.004), plant cover degree (P=0.008), and particle size (P=0.032) comprised the main restrictions of SDX in summer. Soil salinity (P=0.040) and plant height (P=0.011) were the factors influencing CMJ in autumn and winter. This study shows the changing characteristics of macro benthos functional groups to promote coastal wetland restoration and future biogeomorphological studies.
WANG Maoqiu , HU Yang , HE Ning , WU Mingxuan , WU Pengling , WANG Qinyi , ZHANG Bolun , ZHANG Shengle , GAO Meihua , FANG Shubo . The Spatio-temporal Patterns of Macro Benthos Functional Groups and the Associated Factors Affecting Them after Wetland Restoration[J]. Journal of Resources and Ecology, 2022 , 13(6) : 1152 -1164 . DOI: 10.5814/j.issn.1674-764x.2022.06.019
Fig. 1 Research flowchart |
Fig. 2 Study area and sampling sites |
Table 1 The functional group classification of macro benthos |
| Functional groups | Description | Species found in the samples |
|---|---|---|
| CMJ | Jawed mobile carnivore | Palaemon modestus Heller, 1862, Palaemon annandalei Kemp, 1917, Eohaustorius cheliferus Bulyčeva, 1952, Glycera chirori Izuka, 1912, Nephtys glabra Hartman, 1950 |
| CMX | Mobile carnivore | Cerebratulina sp. |
| CDJ | Jawed semi-mobile carnivore | Gnorimosphaeroma rayi Hoestlandt, 1969 |
| FMJ | Jawed mobile suspensivore | Hemileucon bidentatus Liu & Liu, 1990 |
| FMX | Mobile suspensivore | Perioculodes meridichinensis Hirayama, 1992 |
| FDX | Semi-mobile suspensivore | Corbicula fluminea O.F. Müller, 1774; Glaucomya chinensis*, Sinonovacula constricta Lamarck, 1818; Gomphina veneriformis Bivalvia: Veneridae *, Potamocorbula Amurensis Schrenck,1861*; Astarte borealis Schumacher, 1817 |
| SMJ | Jawed mobile surface detritivore | Pyrhila pisum De Haan, 1841; Helicana wuana Rathbun, 1931; Deiratonotus cristatum De Man, 1895; Grandidierella japonica Stephensen, 1938 |
| SDX | Semi-mobile surface detritivore | Assiminea J. Fleming, 1828; Pseudomphala latericea H. Adams & A. Adams, 1864; Assiminea violacea Heude, 1882; Bullacta caurina Benson, 1842; Cerithideopsis largillierti Philippi, 1848 |
| BMJ | Jawed mobile subsurface detritivore | Helice tientsinensis Rathbun, 1931; lyoplax deschampsi Rathbun, 1913 |
| BMX | Tentaculate semi-mobile subsurface detritivore | Heteromastus filiformis Claparède, 1864 |
| HDX | Semi-mobile herbivore | Stenothyra glabra A. Adams, 1861 |
| FSX | Tessile suspensivore | None |
| FST | Tentaculate sessile suspensivore | None |
| SST | Tentaculate sessile surface detritivore | None |
| SMX | Mobile surface detritivore | None |
Note: Feeding habits: F-suspension feeder, S-surface detritivore, B-burrowing detritivore, C-carnivore, H-herbivore. Forms of locomotion: M-mobile, D-semi-mobile, S-sessile. Food gathering methods: J-jawed, T-tentaculate, X-other mechanism. *: Species names were not found on http://www.marinespecies.org, while they appeared in some Chinese sources. |
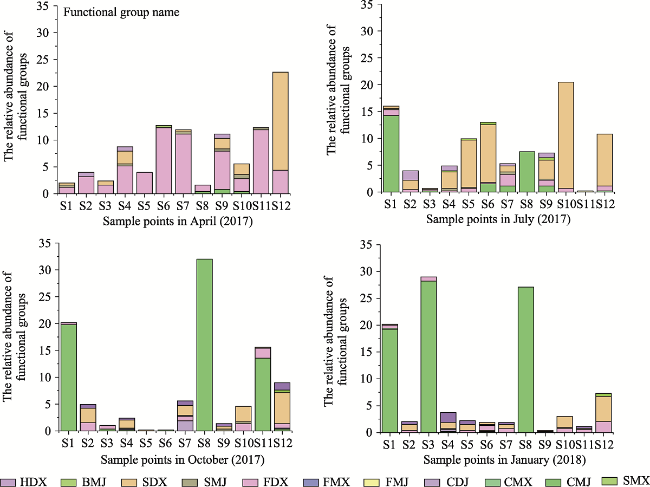
Fig. 3 Changes in the relative abundance of functional groups during the yearNote: FDX dominated in spring. SDX dominated in summer. CMJ dominated in autumn and winter. BMJ, jawed mobile subsurface detritivore; CDJ, jawed semi-mobile carnivore; CMJ, jawed mobile carnivore; CMX, mobile carnivore; FDX, semi-mobile suspensivore; FMJ, jawed mobile suspensivore; FMX, mobile suspensivore; HDX, semi-mobile herbivore; SDX, semi-mobile surface detritivore; SMJ, jawed mobile surface detritivore; SMX, mobile surface detritivore. |
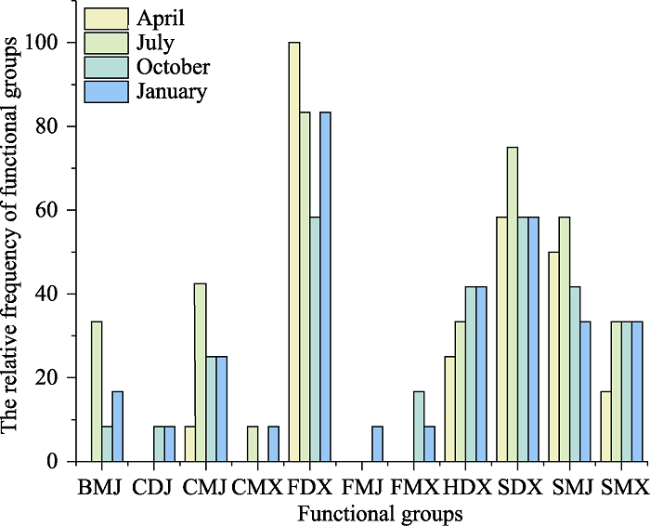
Fig. 4 Changes in the relative frequency of functional groups throughout the yearNote: BMJ: jawed mobile subsurface detritivore; CDJ: jawed semi-mobile carnivore; CMJ: jawed mobile carnivore; CMX: mobile carnivore; FDX: semi-mobile suspensivore; FMJ: jawed mobile suspensivore; FMX: mobile suspensivore; HDX: semi-mobile herbivore; SDX: semi-mobile surface detritivore; SMJ: jawed mobile surface detritivore. SMX: mobile surface detritivore. |
Table 2 The regression analysis of macro benthos functional groups |
| Month | Dependent Variable | Models | Independent Variable | Adjusted R2 | Standard Coefficient B | P |
|---|---|---|---|---|---|---|
| April | CMJ | 1 | EL | 0.849 | ‒0.949 | 0.051 |
| 2 | EL | 0.998 | ‒1.368* | 0.022 | ||
| PD | 0.998 | 0.525 | 0.058 | |||
| FDX | 1 | PZ | 0.376 | ‒0.658* | 0.020 | |
| SMJ | 1 | MC | 0.93 | 0.972** | 0.001 | |
| SDX | 1 | OC | 0.967 | 0.986** | 0.000 | |
| July | CMJ | 1 | PZ | 0.8 | 0.917** | 0.010 |
| 2 | PZ | 0.962 | 1.072** | 0.001 | ||
| TP | 0.962 | ‒0.401* | 0.024 | |||
| SDX | 1 | MC | 0.361 | 0.664 | 0.051 | |
| 2 | MC | 0.59 | 1.158* | 0.011 | ||
| PCD | 0.59 | ‒0.704 | 0.069 | |||
| 3 | MC | 0.821 | 1.085** | 0.004 | ||
| PCD | 0.821 | ‒1.018** | 0.008 | |||
| PZ | 0.821 | ‒0.576* | 0.032 | |||
| BMJ | 1 | PZ | 0.537 | ‒0.793 | 0.060 | |
| HDX | 1 | PH | 0.687 | 0.875 | 0.052 | |
| 2 | PH | 0.948 | 1.143* | 0.013 | ||
| TP | 0.948 | ‒0.531 | 0.057 | |||
| 3 | PH | 0.999 | 1.214* | 0.011 | ||
| TP | 0.999 | ‒0.666* | 0.024 | |||
| MC | 0.999 | 0.198 | 0.069 | |||
| October | CMJ | 1 | PH | 0.708 | ‒0.870* | 0.011 |
| SDX | 1 | PD | 0.363 | 0.685 | 0.090 | |
| BMX | 1 | TP | 0.863 | ‒0.947* | 0.014 | |
| 2 | TP | 0.997 | ‒0.82** | 0.001 | ||
| PZ | 0.997 | ‒0.342** | 0.008 | |||
| HDX | 1 | TP | 0.863 | ‒0.947* | 0.014 | |
| 2 | TP | 0.997 | ‒0.820** | 0.001 | ||
| PZ | 0.997 | ‒0.342** | 0.008 | |||
| January | CMJ | 1 | SS | 0.364 | 0.686 | 0.089 |
| 2 | SS | 0.633 | 0.749* | 0.040 | ||
| TP | 0.633 | 0.538 | 0.097 | |||
| SDX | 1 | MC | 0.695 | 0.864* | 0.012 | |
| BMJ | 1 | MC | . | 1 | . | |
| HDX | 1 | OC | 0.861 | ‒0.946* | 0.015 | |
| FDX | 1 | OC | 0.323 | 0.631* | 0.050 |
Note: EL: elevation, PD: plant density, PZ: particle size, MC: moisture content, OC: organic carbon, TP: total phosphorus, PCD: plant cover degree, SS: soil salinity. |
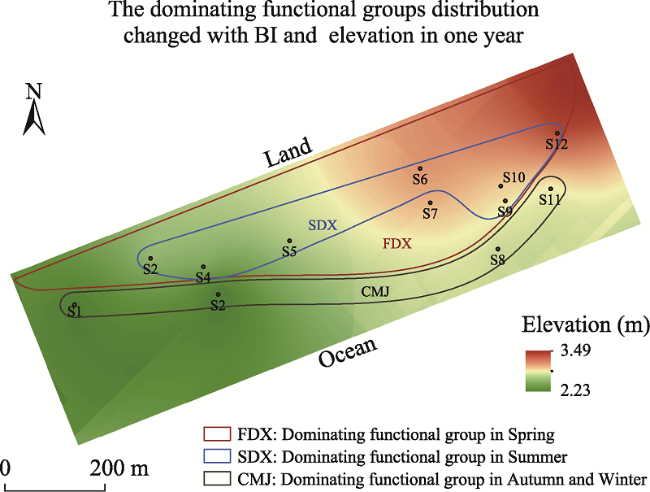
Fig. 5 Distribution of the dominant functional groups in relation to elevation |
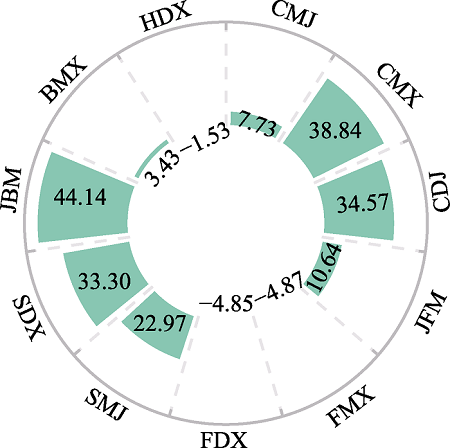
Fig. 6 The BI of all functional groups throughout the year |
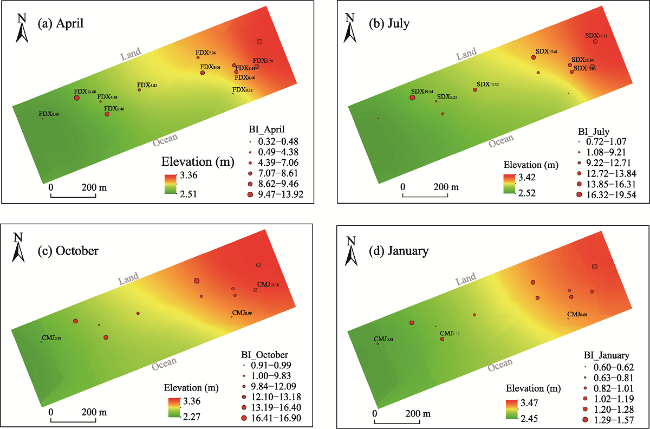
Fig. 7 The change in the distribution of biogeomorphological index (BI) and functional groups throughout the year |
Appendix: The abbreviation list |
| Abbreviation | Full name | Abbreviation | Full name |
|---|---|---|---|
| BI | biogeomorphological index | FST | tentaculate sessile suspensivore |
| BMJ | jawed mobile subsurface detritivore | FSX | tessile suspensivore |
| BMX | tentaculate semi-mobile subsurface detritivore | HDX | semi-mobile herbivore |
| CDJ | jawed semi-mobile carnivore | PCA | principal component analysis |
| CMJ | jawed mobile carnivore | SDX | semi-mobile surface detritivore |
| CMX | mobile carnivore | SMJ | jawed mobile surface detritivore |
| FDX | semi-mobile suspensivore | SMX | mobile surface detritivore |
| FDX | semi-mobile suspensivore | SST | tentaculate sessile surface detritivore |
| FMJ | jawed mobile suspensivore | YRE | the Yangtze River Estuary |
| FMX | mobile suspensivore |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}