
Journal of Resources and Ecology >
How Snow Leopards Share the Same Landscape with Tibetan Agro-pastoral Communities in the Chinese Himalayas
|
XIAO Changxi, E-mail: lzxiaocx@gmail.com |
Received date: 2021-04-16
Accepted date: 2574-07-12
Online published: 2022-04-18
The snow leopard (Panthera uncia) inhabits a human-altered alpine landscape and is often tolerated by residents in regions where the dominant religion is Tibetan Buddhism, including in Qomolangma NNR on the northern side of the Chinese Himalayas. Despite these positive attitudes, many decades of rapid economic development and population growth can cause increasing disturbance to the snow leopards, altering their habitat use patterns and ultimately impacting their conservation. We adopted a dynamic landscape ecology perspective and used multi- scale technique and occupancy model to better understand snow leopard habitat use and coexistence with humans in an 825 km2 communal landscape. We ranked eight hypothetical models containing potential natural and anthropogenic drivers of habitat use and compared them between summer and winter seasons within a year. HABITAT was the optimal model in winter, whereas ANTHROPOGENIC INFLUENCE was the top ranking in summer (AICcw≤2). Overall, model performance was better in the winter than in the summer, suggesting that perhaps some latent summer covariates were not measured. Among the individual variables, terrain ruggedness strongly affected snow leopard habitat use in the winter, but not in the summer. Univariate modeling suggested snow leopards prefer to use rugged land in winter with a broad scale (4000 m focal radius) but with a lesser scale in summer (30 m); Snow leopards preferred habitat with a slope of 22° at a scale of 1000 m throughout both seasons, which is possibly correlated with prey occurrence. Furthermore, all covariates mentioned above showed inextricable ties with human activities (presence of settlements and grazing intensity). Our findings show that multiple sources of anthropogenic activity have complex connections with snow leopard habitat use, even under low human density when anthropogenic activities are sparsely distributed across a vast landscape. This study is also valuable for habitat use research in the future, especially regarding covariate selection for finite sample sizes in inaccessible terrain.
Key words: habitat use; landscape ecology; occupancy model; Qomolangma; Panthera uncia
XIAO Changxi , BAI Defeng , Joseph P. LAMBERT , LI Yibin , Lhaba CERING , GONG Ziling , Philip RIORDAN , SHI Kun . How Snow Leopards Share the Same Landscape with Tibetan Agro-pastoral Communities in the Chinese Himalayas[J]. Journal of Resources and Ecology, 2022 , 13(3) : 483 -500 . DOI: 10.5814/j.issn.1674-764x.2022.03.013
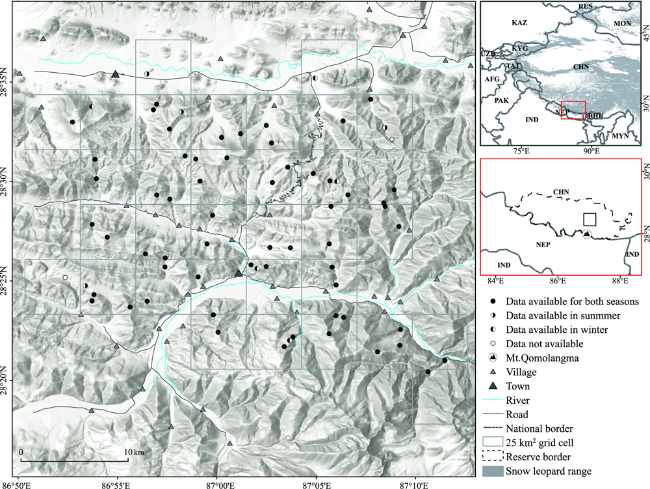
Fig. 1 Location of study area and map of the camera-trap surveyed areas (sites) used to model snow leopard (Panthera uncia) habitat use for summer and winter in Qomolangma NNR, China.Notes: Country names are abbreviated: Afghanistan (AFG); Bhutan (BHU); China (CHN); India (IND); Kazakhstan (KAZ); Kyrgyzstan (KYG); Mongolia (MON); Myanmar (MYN); Nepal (NEP); Pakistan (PAK); Russia (RUS); Tajikistan (TAJ); Uzbekistan (UZB). Projection: UTM45 N; Datum: WGS 1984. |
Table 1 Candidates of snow leopard (Panthera uncia) habitat use (ψ) covariates |
| Category | Covariate | Detail | Abbreviation |
|---|---|---|---|
| Environmental covariates | Blue sheep capture rates | Camera-trap capture | BS |
| Sites of slope gradient | 30 m, 300 m, 1000 m, 2000 m, 4000 m | SLP30-4000 | |
| Sites of terrain ruggedness index | 30 m, 300 m, 1000 m, 2000 m, 4000 m | TRI30-4000 | |
| Sites of elevation | 30 m, 300 m, 1000 m, 2000 m, 4000 m | ELE30-4000 | |
| Anthropogenic covariates | Grazing activity rates | Camera-trap capture | GRAZE |
| Disturbance from human settlements | Kernal density | KD_SETT | |
| Distance of the nearest | Dist._SETT | ||
| Disturbance from traffic roads | Kernal density | KD_ROAD | |
| Distance of the nearest | Dist._ROAD |
Table 2 Description of priori candidate models describing the effects of potential environmental variables on the probability of habitat use by snow leopard (Panthera uncia) |
| Model name (Abbreviation) | Covariates (Abbreviation) | Hypothesis | Expected influence on ψ | Supporting literature | |
|---|---|---|---|---|---|
| 1 | NULL | N/A | Snow leopard habitat use is not affected by environmental variables | N/A | N/A |
| 2 | PREY | Blue sheep capture rates (BS) | Snow leopards use habitat based on access to prey | Positive | Barber-Meyer et al., 2013; McCarthy, 2000; Sharma et al., 2015 |
| 3 | TOPOGRAPHY (TOP.) | Slope (SLP) | Snow leopards use high mountain areas with rugged and steep slope terrain | Positive | Sunarto et al., 2012; Taubmann et al., 2016; Klaassen and Broekhuis, 2018; Ghoshal et al., 2019; Watts et al., 2019 |
| Terrain ruggedness index (TRI) | Positive | ||||
| Elevation (ELE) | Positive | ||||
| 4 | HABITAT | Blue sheep capture rates (BS) | Snow leopards use high, steep slope, and rugged habitat where prey is abundant | Positive | Sunarto et al., 2012; Barber-Meyer et al., 2013; Taubmann et al., 2016; Klaassen and Broekhuis, 2018; Ghoshal et al., 2019 |
| All potential TOP. variables | Positive | ||||
| 5 | ANTHROPOGENIC INFLUENCE (A.I.) | Grazing activity index (GRAZE) | Snow leopards avoid areas with high levels of human disturbance | Negative | McCarthy and Chapron, 2003; Cosentino et al., 2014; Lewis et al., 2015; Alexander et al., 2016a |
| Disturbance from human settlements (SETT) | Negative | ||||
| Disturbance from traffic roads (ROAD) | Negative | ||||
| 6 | PREY+A.I. | Blue sheep capture rates (BS) | Snow leopards use habitat based on access to prey and avoiding human disturbance | Positive | McCarthy, 2000; McCarthy and Chapron, 2003; Alexander et al., 2016a |
| All potential A.I. variables | Negative | ||||
| 7 | TOP.+A.I. | All potential TOP. variables | Snow leopards use rugged, high elevation habitat with little human disturbance | Positive | Sunarto et al., 2012; Alexander et al., 2016a; Klaassen and Broekhuis, 2018; Ghoshal et al., 2019 |
| All potential A.I. variables | Negative | ||||
| 8 | GLOBAL | Blue sheep capture rates (BS) | Snow leopards use high elevation, rugged habitat where prey is abundant and with low human disturbance | Positive | McCarthy and Chapron, 2003; Sunarto et al., 2012; Taubmann et al., 2016; Ghoshal et al., 2019 |
| All potential TOP. variables | Positive | ||||
| All potential A.I. variables | Negative |
Note: All models are based on hypotheses developed from the cited supporting literature. The models shown here are before removing correlated variables, and those which were non-optimum candidates at the stage of the univariate habitat modeling and true multi-scale evaluation. For the full model set included in the final analysis, see Table 3. No paremeters (N/A). |
Table 3 Description of a priori candidate models describing the effects of environmental variables on the probability of habitat use by snow leopard (Panthera uncia) |
| Covariate (unit) | Abbreviation (scale) | Relationship to snow leopard occurrence | Season | Effective sample range (mean±SD) | Station ratio recorded to target | Supporting literature |
|---|---|---|---|---|---|---|
| Grazing activity capture rate index (%) | GRAZE (N/A) | Avoids sites with high levels of human disturbance | Winter | 0-54.55 (7.59±10.24) | 88.53 | McCarthy and Chapron, 2003; Alexander et al., 2016a |
| Summer | 0-67.78 (14.71±16.44) | 95.16 | ||||
| Blue sheep capture rate index (%) | BS (N/A) | Habitat use based on access to prey | Winter | 0-36.67 (2.80±5.6) | 47.54 | McCarthy, 2000; Barber-Meyer et al., 2013; Sharma et al., 2015 |
| Summer | 0-20.45 (2.14±3.66) | 50.77 | ||||
| Slope of sites (°) | SLP (1000) | Uses certain steep habitat | Winter | 15.93-32.70 (22.15±3.17) | N/A | Sunarto et al., 2012; Klaassen and Broekhuis, 2018 |
| SLP (1000) | Summer | 7.49-32.70 (22.03±3.77) | ||||
| Terrain ruggedness index (N/A) | TRI (4000) | Uses rugged terrain habitat | Winter | 12.11-21.36 (15.50±1.96) | N/A | Taubmann et al., 2016; Ghoshal et al., 2019 |
| TRI (30) | Summer | 1.40-4.09 (2.34±0.57) |
Note: All models are based on hypotheses developed from the cited supporting literature. No paremeters (N/A). |
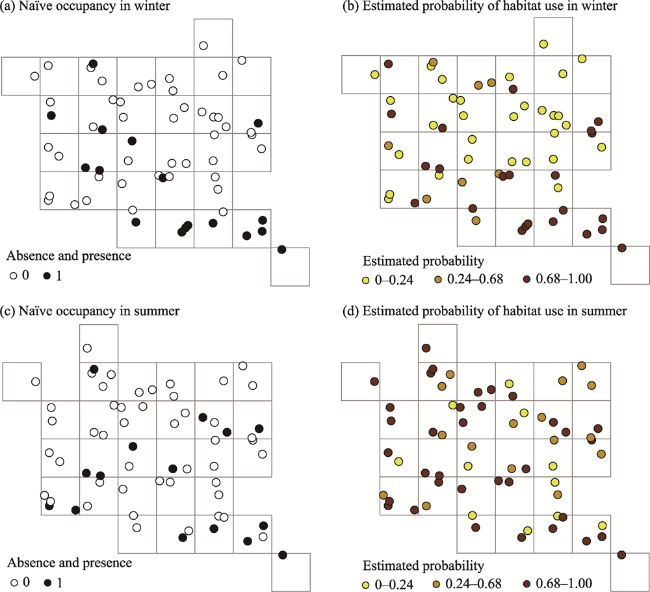
Fig. 2 Probability of site use by snow leopards (Panthera uncia), as measured by camera-traps in Qomolangma NNR, China. (a, c) Naïve estimates from a presence vs. absence approach of two seasons; (b, d) Mean estimated probabilities of habitat use of two seasons.Note: Each cell is 5 km×5 km=25 km2. |
Table 4 Snow leopard (Panthera uncia) detection probability models (p) in winter and summer |
| Models_winter | AICc | ∆AICc | AICcw | Model likelihood | K | -2 log LL |
|---|---|---|---|---|---|---|
| p(TEAM) | 185.15 | 0 | 0.7917 | 1 | 3 | 178.73 |
| p(.) | 187.82 | 2.67 | 0.2083 | 0.2632 | 2 | 183.61 |
| Models_summer | AICc | ∆AICc | AICcw | Model likelihood | K | -2 log LL |
| p(TEAM) | 150.5 | 0 | 0.6857 | 1 | 3 | 144.09 |
| p(.) | 152.06 | 1.56 | 0.3143 | 0.4584 | 2 | 147.86 |
Note: Akaike's information criterion corrected for finite sample sizes (AICc). The relative difference in AICc values compared with the top-ranked model (ΔAICc), model weight (AICcw), Model likelihood, number of parameters (K), and -2log-likelihood (-2 log LL). The detection covariate was observer bias of camera station setting teams (TEAM). The dataset was limited to 90 sampling days with a 15-day collapsing scenario, 61 valid camera-trap stations in winter, and an 18-day collapsing scenario, 62 valid camera-trap stations in summer. |
Table 5 Model selection results for describing the probability of snow leopard (Panthera uncia) site use (ψ) in the cold and warm seasons |
| No. | Models_winter | AICc | ∆AICc | AICcw | Model likelihood | K | -2 log LL |
|---|---|---|---|---|---|---|---|
| 1 | HABITAT | 162.26 | 0 | 0.4155 | 1 | 6 | 148.7 |
| 2 | TOP. | 162.61 | 0.35 | 0.3488 | 0.8395 | 5 | 151.52 |
| 3 | GLOBAL | 164.55 | 2.29 | 0.1322 | 0.3182 | 7 | 148.44 |
| 4 | TOP. +A.I. | 165.04 | 2.78 | 0.1035 | 0.2491 | 6 | 151.48 |
| 5 | A.I. | 187.16 | 24.9 | 0 | 0 | 4 | 178.45 |
| 6 | PREY | 187.26 | 25 | 0 | 0 | 4 | 178.55 |
| 7 | NULL | 187.82 | 25.56 | 0 | 0 | 2 | 183.61 |
| 8 | PREY+A.I. | 189.32 | 27.06 | 0 | 0 | 5 | 178.23 |
| No. | Models_summer | AICc | ∆AICc | AICcw | Model likelihood | K | -2 log LL |
| 1 | A.I. | 150.57 | 0 | 0.2152 | 1 | 4 | 141.87 |
| 2 | TOP. | 150.68 | 0.11 | 0.2037 | 0.9465 | 5 | 139.61 |
| 3 | PREY+A.I. | 151.59 | 1.02 | 0.1292 | 0.6005 | 5 | 140.52 |
| 4 | PREY | 151.67 | 1.10 | 0.1242 | 0.5769 | 4 | 142.97 |
| 5 | TOP. +A.I. | 151.84 | 1.27 | 0.1141 | 0.5299 | 6 | 138.31 |
| 6 | NULL | 152.06 | 1.49 | 0.1022 | 0.4747 | 2 | 147.86 |
| 7 | HABITAT | 152.70 | 2.13 | 0.0742 | 0.3447 | 6 | 139.17 |
| 8 | GLOBAL | 154.08 | 3.51 | 0.0372 | 0.1729 | 7 | 138.01 |
Note: Akaike's information criterion corrected for finite sample sizes (AICc). The relative difference in AICc values compared with the top-ranked model (ΔAICc), model weight (AICcw), Model likelihood, number of parameters (K), and -2log-likelihood (-2 log LL). |
Table 6 Estimates of the Beta coefficient (β) and standard error (SE) of each covariate of the hypothetical models |
| No. | Models_winter | βintercepte ±SE | βGRAZE±SE | βBS±SE | βSLP1000±SE | βTRI30±SE | βTRI4000±SE |
|---|---|---|---|---|---|---|---|
| 1 | HABITAT | 0.38±0.99 | N/A | -1.79±1.74 | 7.41±4.73 | N/A | 2.44±1.41 |
| 2 | TOP. | -0.27±0.75 | N/A | N/A | 3.89±2.06 | N/A | 1.62±1.01 |
| 3 | GLOBAL | 0.32±0.95 | -0.55±1.21 | -1.69±1.62 | 6.68±4.24 | N/A | 2.44±1.36 |
| 4 | TOP. +A.I. | -0.26±0.76 | -0.15±0.74 | N/A | 3.90±2.04 | N/A | 1.66±1.04 |
| 5 | A.I. | -0.19±0.45 | -0.21±0.40 | N/A | N/A | N/A | N/A |
| 6 | PREY | -0.18±0.45 | N/A | -0.15±0.36 | N/A | N/A | N/A |
| 7 | NULL | -0.60±0.36 | N/A | N/A | N/A | N/A | N/A |
| 8 | PREY+A.I. | -0.18±0.45 | -0.23±0.41 | -0.17±0.36 | N/A | N/A | N/A |
| No. | Models_summer | βintercepte ±SE | βGRAZE±SE | βBS±SE | βSLP1000±SE | βTRI30±SE | βTRI4000±SE |
| 1 | A.I. | -0.34±0.55 | -0.77±0.57 | N/A | N/A | N/A | N/A |
| 2 | TOP. | -0.20±0.58 | N/A | N/A | 0.65±0.57 | 0.51±0.52 | N/A |
| 3 | PREY+A.I. | 0.17±1.25 | -0.97±0.89 | 1.58±2.36 | N/A | N/A | N/A |
| 4 | PREY | 0.06±0.72 | N/A | 0.94±1.03 | N/A | N/A | N/A |
| 5 | TOP. +A.I. | -0.38±0.56 | -0.63±0.54 | N/A | 0.52±0.48 | 0.49±0.52 | N/A |
| 6 | NULL | -0.57±0.44 | N/A | N/A | N/A | N/A | N/A |
| 7 | HABITAT | -0.12±0.65 | N/A | -0.37±0.57 | 0.80±0.66 | 0.70±0.66 | N/A |
| 8 | GLOBAL | -0.35±0.59 | -0.64±0.56 | -0.29±0.51 | 0.59±0.50 | 0.63±0.59 | N/A |
Note: Grazing activity index (GRAZE), blue sheep capture rate (BS), slope steepness of sites with scale 1000 m threshold value (SLP1000), terrain ruggedness index with scale 30 m and 4000 m threshold values (TRI30, TRI4000). No paremeters (N/A). |
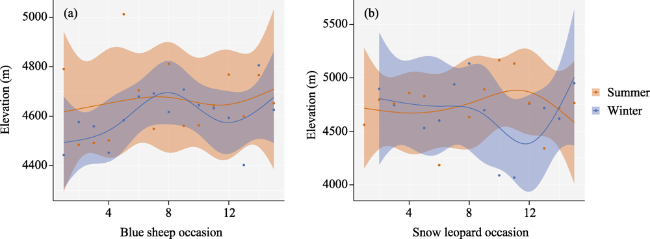
Fig. 3 Seasonal variation in elevations for blue sheep (Pseduois nayaur) and snow leopard (Panthera uncia) in two seasons, winter and summer.Note: Lines show medians of the posterior distribution, shaded areas show the 95% confidence intervals, and points show each of the 6-day means of the raw data for 90 days of winter (Nov. 2017-Jan. 2018) and summer (May-Jul. 2018). |
Table S1 Chi-square probability (χ2p) and over dispersion statistic c-hat (ĉ) results of the MacKenzie and Bailey (2004) goodness-of-fit (GoF) test for snow leopard (Panthera uncia) habitat use models with different collapsing day-periods |
| Collapsing scenarios_winter | χ²p | ĉ | Collapsing scenarios_summer | χ²p | ĉ |
|---|---|---|---|---|---|
| 5-day sampling occasions | 0.1882 | 0.4276 | 5-day sampling occasions | 0.0133 | 7.4845 |
| 10-day sampling occasions | 0.1414 | 1.3309 | 10-day sampling occasions | 0.0782 | 1.9145 |
| 15-day sampling occasions | 0.2261 | 1.0823 | 15-day sampling occasions | 0.1551 | 1.2427 |
| 18-day sampling occasions | 0.1566 | 1.2545 | 18-day sampling occasions | 0.2545 | 1.0112 |
Note: Detection probability (p) was a binary covariate indicating which of the two survey teams placed a particular camera trap (TEAM). Site use covariates were blue sheep camera-trap capture rates (BS), grazing activity camera trap rates (GRAZE), terrain ruggedness index (TRI) with multiple scales of 30 m, 300 m, 1000 m, 2000 m, and 4000 m (TRI30, TRI300, TRI1000, TRI2000, TRI4000), slopes (SLP) with multi-scales of 30 m, 300 m, 1000 m, 2000 m, 4000 m (SLP30, SLP300, SLP1000, SLP2000, SLP4000). The dataset was limited to 90 sampling days in each season and the effective camera-trap stations numbered 61 and 62 in winter and summer, respectively. |
Table S2 Summary of model-averaged parameter estimates of the probability of habitat use (ψ) and detection (p̂) for snow leopards (Panthera uncia) at two seasons in Qomolangma NNR |
| Models_winter | ψ±SE | p̂±SE | Models_summer | ψ±SE | p̂±SE |
|---|---|---|---|---|---|
| HABITAT | 0.46±0.08 | 0.18±0.04 | A.I. | 0.44±0.15 | 0.18±0.06 |
| TOP. | 0.43±0.09 | 0.19±0.05 | TOP. | 0.45±0.18 | 0.18±0.06 |
| GLOBAL | 0.46±0.09 | 0.18±0.04 | PREY+A.I. | 0.50±0.22 | 0.16±0.06 |
| TOP. +A.I. | 0.43±0.10 | 0.19±0.05 | PREY | 0.49±0.17 | 0.17±0.06 |
| A.I. | 0.45±0.13 | 0.18±0.05 | TOP. +A.I. | 0.42±0.18 | 0.19±0.06 |
| PREY | 0.46±0.13 | 0.18±0.05 | NULL | 0.36±0.10 | 0.23±0.07 |
| NULL | 0.35±0.08 | 0.24±0.05 | HABITAT | 0.47±0.20 | 0.18±0.06 |
| PREY+A.I. | 0.46±0.15 | 0.18±0.06 | GLOBAL | 0.43±0.20 | 0.19±0.06 |
| Model averaged | 0.44±0.11 | 0.19±0.05 | Model averaged | 0.45±0.18 | 0.19±0.06 |
Note: Models: TOPOGRAPHY (TOP.), ANTHROPOGENIC INFLUENCE (A.I.); standard error (SE); model-averaged estimates across all models with unconditional standard errors. |
Table S3 Correlation matrix of the continuous covariates in winter |
| Dist._ sett | KD_ sett | Dist._ road | KD_ road | TRI30 | TRI300 | TRI1000 | TRI2000 | TRI4000 | ELE30 | ELE300 | ELE1000 | ELE2000 | ELE4000 | BS | Graze | SLP30 | SLP300 | SLP1000 | SLP2000 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KD_sett | -0.632 | |||||||||||||||||||
| Dist._road | 0.233 | -0.443 | ||||||||||||||||||
| KD_road | 0.071 | 0.188 | -0.645 | |||||||||||||||||
| TRI30 | -0.064 | 0.127 | -0.141 | -0.066 | ||||||||||||||||
| TRI300 | -0.186 | 0.297 | -0.126 | -0.092 | 0.441 | |||||||||||||||
| TRI1000 | -0.324 | 0.372 | -0.120 | -0.112 | 0.330 | 0.657 | ||||||||||||||
| TRI2000 | -0.336 | 0.305 | -0.143 | -0.090 | 0.242 | 0.436 | 0.832 | |||||||||||||
| TRI4000 | -0.148 | -0.015 | 0.091 | -0.220 | 0.216 | 0.398 | 0.615 | 0.775 | ||||||||||||
| ELE30 | 0.549 | -0.475 | 0.385 | 0.030 | -0.263 | -0.262 | -0.333 | -0.321 | -0.149 | |||||||||||
| ELE300 | 0.562 | -0.513 | 0.396 | 0.027 | -0.257 | -0.303 | -0.359 | -0.346 | -0.169 | 0.995 | ||||||||||
| ELE1000 | 0.615 | -0.600 | 0.413 | 0.009 | -0.233 | -0.352 | -0.394 | -0.402 | -0.242 | 0.944 | 0.968 | |||||||||
| ELE2000 | 0.643 | -0.699 | 0.445 | 0.000 | -0.240 | -0.359 | -0.420 | -0.395 | -0.229 | 0.860 | 0.892 | 0.959 | ||||||||
| ELE4000 | 0.632 | -0.796 | 0.478 | -0.036 | -0.164 | -0.244 | -0.377 | -0.342 | -0.120 | 0.746 | 0.780 | 0.851 | 0.940 | |||||||
| BS | -0.346 | 0.142 | -0.105 | -0.153 | 0.158 | 0.159 | 0.336 | 0.283 | 0.109 | -0.192 | -0.190 | -0.168 | -0.171 | -0.154 | ||||||
| Graze | 0.094 | -0.194 | 0.098 | -0.071 | 0.007 | -0.160 | -0.067 | -0.032 | -0.049 | 0.001 | 0.026 | 0.087 | 0.152 | 0.157 | -0.063 | |||||
| SLP30 | -0.172 | 0.287 | -0.259 | -0.035 | 0.911 | 0.563 | 0.374 | 0.318 | 0.262 | -0.385 | -0.394 | -0.404 | -0.425 | -0.344 | 0.181 | -0.080 | ||||
| SLP300 | -0.151 | 0.284 | -0.178 | -0.085 | 0.326 | 0.896 | 0.622 | 0.438 | 0.406 | -0.213 | -0.253 | -0.323 | -0.335 | -0.240 | 0.137 | -0.170 | 0.443 | |||
| SLP1000 | -0.027 | 0.142 | 0.004 | -0.220 | 0.272 | 0.582 | 0.691 | 0.554 | 0.557 | -0.138 | -0.160 | -0.192 | -0.232 | -0.187 | 0.143 | -0.171 | 0.260 | 0.711 | ||
| SLP2000 | 0.072 | -0.202 | 0.234 | -0.323 | 0.262 | 0.366 | 0.513 | 0.500 | 0.703 | -0.052 | -0.055 | -0.071 | -0.048 | 0.047 | 0.135 | -0.131 | 0.215 | 0.468 | 0.809 | |
| SLP4000 | 0.177 | -0.394 | 0.231 | -0.200 | 0.232 | 0.236 | 0.359 | 0.372 | 0.623 | -0.066 | -0.054 | -0.061 | 0.015 | 0.205 | 0.023 | -0.074 | 0.164 | 0.302 | 0.519 | 0.831 |
Note: Pairs of covariates were considered as having high collinearity when |r|>0.6 (values in bold). Site covariates tested were: elevation with five multi-scale 30 m, 300 m, 1000 m, 2000 m, 4000 m threshold values (ELE30, ELE300, ELE1000, ELE2000, ELE4000), slope with five multi-scale threshold values 30 m, 300 m, 1000 m, 2000 m, 4000 m (SLP30, SLP300, SLP1000, SLP2000, SLP4000), terrain ruggedness index with five multi-scale threshold values 30 m, 300 m, 1000 m, 2000 m, 4000 m (TRI30, TRI300, TRI1000, TRI2000,TRI4000), grazing activity index (GRAZE), blue sheep index (BS), kernel density of human settlement (KD_SETT) and road (KD_ROAD), and distance to the human settlement (Dist._SETT) and road (Dist._ROAD). |
Table S4 Correlation matrix of the continuous covariates in summer |
| BS | Graze | Dist._ SETT | KD_ SETT | Dist._ ROAD | KD_ ROAD | TRI30 | TRI300 | TRI1000 | TRI2000 | TRI4000 | ELE30 | ELE300 | ELE1000 | ELE2000 | ELE4000 | SLP30 | SLP300 | SLP1000 | SLP2000 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Graze | -0.093 | |||||||||||||||||||
| Dist._SETT | -0.109 | -0.126 | ||||||||||||||||||
| KD_SETT | 0.002 | 0.051 | -0.629 | |||||||||||||||||
| Dist._ROAD | -0.018 | -0.115 | 0.290 | -0.492 | ||||||||||||||||
| KD_ROAD | -0.124 | 0.140 | 0.030 | 0.205 | -0.631 | |||||||||||||||
| TRI30 | 0.184 | 0.020 | -0.151 | 0.141 | -0.175 | -0.072 | ||||||||||||||
| TRI300 | 0.332 | -0.044 | -0.149 | 0.170 | -0.160 | -0.062 | 0.434 | |||||||||||||
| TRI1000 | 0.327 | 0.012 | -0.209 | 0.161 | -0.060 | -0.086 | 0.320 | 0.670 | ||||||||||||
| TRI2000 | 0.301 | 0.072 | -0.229 | 0.160 | -0.118 | -0.039 | 0.275 | 0.481 | 0.865 | |||||||||||
| TRI4000 | 0.399 | 0.059 | -0.088 | -0.058 | 0.028 | -0.125 | 0.275 | 0.432 | 0.665 | 0.793 | ||||||||||
| ELE30 | -0.136 | -0.243 | 0.581 | -0.543 | 0.422 | 0.005 | -0.362 | -0.319 | -0.245 | -0.246 | -0.153 | |||||||||
| ELE300 | -0.159 | -0.214 | 0.587 | -0.576 | 0.434 | 0.001 | -0.363 | -0.348 | -0.256 | -0.258 | -0.166 | 0.995 | ||||||||
| ELE1000 | -0.203 | -0.150 | 0.626 | -0.649 | 0.454 | -0.018 | -0.340 | -0.355 | -0.253 | -0.277 | -0.218 | 0.950 | 0.971 | |||||||
| ELE2000 | -0.188 | -0.125 | 0.642 | -0.732 | 0.482 | -0.019 | -0.347 | -0.340 | -0.259 | -0.273 | -0.219 | 0.874 | 0.901 | 0.960 | ||||||
| ELE4000 | -0.148 | -0.124 | 0.627 | -0.818 | 0.502 | -0.042 | -0.262 | -0.221 | -0.198 | -0.211 | -0.112 | 0.752 | 0.782 | 0.851 | 0.942 | |||||
| SLP30 | 0.211 | 0.023 | -0.228 | 0.312 | -0.295 | -0.027 | 0.908 | 0.548 | 0.347 | 0.326 | 0.300 | -0.462 | -0.475 | -0.480 | -0.496 | -0.410 | ||||
| SLP300 | 0.420 | -0.036 | -0.103 | 0.158 | -0.215 | -0.057 | 0.279 | 0.871 | 0.627 | 0.478 | 0.439 | -0.278 | -0.301 | -0.316 | -0.310 | -0.217 | 0.399 | |||
| SLP1000 | 0.413 | -0.062 | 0.023 | -0.024 | 0.026 | -0.177 | 0.191 | 0.586 | 0.741 | 0.649 | 0.620 | -0.115 | -0.120 | -0.101 | -0.129 | -0.079 | 0.189 | 0.734 | ||
| SLP2000 | 0.483 | -0.026 | 0.083 | -0.239 | 0.204 | -0.252 | 0.200 | 0.438 | 0.662 | 0.635 | 0.733 | -0.040 | -0.036 | -0.020 | -0.009 | 0.073 | 0.167 | 0.553 | 0.884 | |
| SLP4000 | 0.390 | -0.018 | 0.168 | -0.372 | 0.203 | -0.158 | 0.210 | 0.332 | 0.565 | 0.551 | 0.695 | -0.078 | -0.064 | -0.043 | 0.025 | 0.194 | 0.153 | 0.409 | 0.666 | 0.867 |
Note: Pairs of covariates were considered as having high collinearity when |r|>0.6 (values in bold). Site covariates tested were: elevation with five multi-scale threshold values 30 m, 300 m, 1000 m, 2000 m, 4000 m (ELE30, ELE300, ELE1000, ELE2000, ELE4000), slope with five multi-scale threshold values 30 m, 300 m, 1000 m, 2000 m, 4000 m (SLP30, SLP300, SLP1000, SLP2000, SLP4000), terrain ruggedness index with five multi-scale threshold values 30 m, 300 m, 1000 m, 2000 m, 4000 m (TRI30, TRI300, TRI1000, TRI2000,TRI4000), grazing activity index (GRAZE), blue sheep index (BS), kernel density of human settelment (KD_SETT) and road (KD_ROAD), and distance to the human settlement (Dist._SETT) and road (Dist._ROAD). |
Table S5 Portion of snow leopard (Panthera uncia) single covariate occupancy models (ψ) in winter and summer |
| Models_winter | AICc | ∆AICc | AICcw | Model Likelihood | K | -2 log LL | Models_summer | AICc | ∆AICc | AICcw | Model Likelihood | K | -2 log LL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ψ(SLP1000) | 165.17 | 0.00 | 0.9517 | 1.0000 | 4 | 156.46 | ψ(SLP1000) | 149.41 | 0.00 | 0.1154 | 1.0000 | 4 | 140.71 |
| ψ(TRI4000) | 173.10 | 7.93 | 0.0181 | 0.0190 | 4 | 164.39 | ψ(SLP4000) | 150.07 | 0.66 | 0.0830 | 0.7189 | 4 | 141.37 |
| ψ(TRI1000) | 173.17 | 8.00 | 0.0174 | 0.0183 | 4 | 164.46 | ψ(TRI30) | 150.16 | 0.75 | 0.0793 | 0.6873 | 4 | 141.46 |
| ψ(SLP300) | 176.19 | 11.02 | 0.0039 | 0.0040 | 4 | 167.48 | ψ(SLP2000) | 150.21 | 0.80 | 0.0774 | 0.6703 | 4 | 141.51 |
| ψ(SLP2000) | 176.49 | 11.32 | 0.0033 | 0.0035 | 4 | 167.78 | ψ(.) | 150.50 | 1.09 | 0.0669 | 0.5798 | 3 | 144.09 |
| ψ(TRI2000) | 176.67 | 11.50 | 0.0030 | 0.0032 | 4 | 167.96 | ψ(GRAZE) | 150.57 | 1.16 | 0.0646 | 0.5599 | 4 | 141.87 |
| ψ(TRI300) | 177.94 | 12.77 | 0.0016 | 0.0017 | 4 | 169.23 | ψ(TRI4000) | 150.80 | 1.39 | 0.0576 | 0.4991 | 4 | 142.10 |
| ψ(ELE2000) | 181.79 | 16.62 | 0.0002 | 0.0002 | 4 | 173.08 | ψ(SLP300) | 150.87 | 1.46 | 0.0556 | 0.4819 | 4 | 142.17 |
| ψ(SLP4000) | 182.43 | 17.26 | 0.0002 | 0.0002 | 4 | 173.72 | ψ(SLP30) | 150.91 | 1.50 | 0.0545 | 0.4724 | 4 | 142.21 |
| ψ(ELE4000) | 183.20 | 18.03 | 0.0001 | 0.0001 | 4 | 174.49 | ψ(TRI1000) | 151.47 | 2.06 | 0.0412 | 0.3570 | 4 | 142.77 |
| ψ(ELE1000) | 183.51 | 18.34 | 0.0001 | 0.0001 | 4 | 174.80 | ψ(BS) | 151.67 | 2.26 | 0.0373 | 0.3230 | 4 | 142.97 |
| ψ(KD_SETT) | 184.08 | 18.91 | 0.0001 | 0.0001 | 4 | 175.37 | ψ(TRI300) | 152.02 | 2.61 | 0.0313 | 0.2712 | 4 | 143.32 |
| ψ(.) | 185.15 | 19.98 | 0.0000 | 0.0000 | 3 | 178.73 | ψ(Dist._SETT) | 152.20 | 2.79 | 0.0286 | 0.2478 | 4 | 143.50 |
| ψ(ELE300) | 185.34 | 20.17 | 0.0000 | 0.0000 | 4 | 176.63 | ψ(KD_ROAD) | 152.41 | 3.00 | 0.0258 | 0.2231 | 4 | 143.71 |
| ψ(SLP30) | 185.58 | 20.41 | 0.0000 | 0.0000 | 4 | 176.87 | ψ(TRI2000) | 152.43 | 3.02 | 0.0255 | 0.2209 | 4 | 143.73 |
| ψ(TRI30) | 185.92 | 20.75 | 0.0000 | 0.0000 | 4 | 177.21 | ψ(Dist._ROAD) | 152.52 | 3.11 | 0.0244 | 0.2112 | 4 | 143.82 |
| ψ(ELE30) | 185.96 | 20.79 | 0.0000 | 0.0000 | 4 | 177.25 | ψ(ELE2000) | 152.64 | 3.23 | 0.0230 | 0.1989 | 4 | 143.94 |
| ψ(KD_ROAD) | 186.06 | 20.89 | 0.0000 | 0.0000 | 4 | 177.35 | ψ(ELE4000) | 152.71 | 3.30 | 0.0222 | 0.1920 | 4 | 144.01 |
| ψ(Dist._SETT) | 186.40 | 21.23 | 0.0000 | 0.0000 | 4 | 177.69 | ψ(ELE30) | 152.75 | 3.34 | 0.0217 | 0.1882 | 4 | 144.05 |
| ψ(GRAZE) | 187.16 | 21.99 | 0.0000 | 0.0000 | 4 | 178.45 | ψ(ELE300) | 152.76 | 3.35 | 0.0216 | 0.1873 | 4 | 144.06 |
| ψ(BS) | 187.26 | 22.09 | 0.0000 | 0.0000 | 4 | 178.55 | ψ(KD_SETT) | 152.77 | 3.36 | 0.0215 | 0.1864 | 4 | 144.07 |
| ψ(Dist._ROAD) | 187.37 | 22.20 | 0.0000 | 0.0000 | 4 | 178.66 | ψ(ELE1000) | 152.78 | 3.37 | 0.0214 | 0.1854 | 4 | 144.08 |
Note: Akaike's information criterion corrected for small sample sizes (AICc), relative difference in AICc values compared with the top ranked model (ΔAICc), model weight (AICcw), Model Likelihood, number of parameters (K), and -2log-likelihood (-2 log LL). Detection covariate was camera trap deployment team (TEAM). Site covariates were: elevation with multi-scale threshold values of 30 m, 300 m, 1000 m, 2000 m, 4000 m (ELE30, ELE300, ELE1000, ELE2000, ELE4000), slope with multi-scale threshold values of 30 m, 300 m, 1000 m, 2000 m, 4000 m (SLP30, SLP300, SLP1000, SLP2000, SLP4000), terrain ruggedness index with multi-scale threshold values of 30 m, 300 m, 1000 m, 2000 m, 4000 m (TRI30, TRI300, TRI1000, TRI2000,TRI4000), grazing activity index (GRAZE), blue sheep index (BS), kernel density of human settlement (KD_SETT) and road (KD_ROAD), and distance to the human settlement (Dist._SETT) and road (Dist._ROAD). |
Table S6 True optimum multi-scale approach of topographic gradient variables of snow leopard (Panthera uncia) habitat use in winter and summer |
| Models_winter | AICc | ∆AICc | AICcw | Model Likelihood | K | -2 log LL | Models_summer | AICc | ∆AICc | AICcw | Model Likelihood | K | -2 log LL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ψ(TRI4000+SLP1000) | 162.61 | 0.00 | 0.7130 | 1.0000 | 5 | 151.52 | ψ(.) | 150.50 | 0.00 | 0.1259 | 1.0000 | 3 | 144.09 |
| ψ(TRI2000+SLP1000) | 166.46 | 3.85 | 0.1040 | 0.1459 | 5 | 155.37 | ψ(TRI30+SLP1000) | 150.68 | 0.18 | 0.1151 | 0.9139 | 5 | 139.61 |
| ψ(TRI300+SLP1000) | 167.09 | 4.48 | 0.0759 | 0.1065 | 5 | 156.00 | ψ(TRI30+SLP4000) | 150.99 | 0.49 | 0.0985 | 0.7827 | 5 | 139.92 |
| ψ(TRI30+SLP1000) | 167.49 | 4.88 | 0.0621 | 0.0872 | 5 | 156.40 | ψ(TRI30+SLP2000) | 151.34 | 0.84 | 0.0827 | 0.6570 | 5 | 140.27 |
| ψ(TRI4000+SLP300) | 169.02 | 6.41 | 0.0289 | 0.0406 | 5 | 157.93 | ψ(TRI300+SLP1000) | 151.71 | 1.21 | 0.0688 | 0.5461 | 5 | 140.64 |
| ψ(TRI1000+SLP4000) | 173.17 | 10.56 | 0.0036 | 0.0051 | 5 | 162.08 | ψ(TRI30+SLP300) | 151.95 | 1.45 | 0.0610 | 0.4843 | 5 | 140.88 |
| ψ(TRI300+SLP2000) | 173.29 | 10.68 | 0.0034 | 0.0048 | 5 | 162.20 | ψ(TRI2000+SLP4000) | 151.95 | 1.45 | 0.0610 | 0.4843 | 5 | 140.88 |
| ψ(TRI2000+SLP300) | 174.19 | 11.58 | 0.0022 | 0.0031 | 5 | 163.10 | ψ(TRI4000+SLP30) | 152.05 | 1.55 | 0.0580 | 0.4607 | 5 | 140.98 |
| ψ(TRI1000+SLP2000) | 174.40 | 11.79 | 0.0020 | 0.0028 | 5 | 163.31 | ψ(TRI300+SLP4000) | 152.39 | 1.89 | 0.0489 | 0.3887 | 5 | 141.32 |
| ψ(TRI1000+SLP30) | 175.21 | 12.60 | 0.0013 | 0.0018 | 5 | 164.12 | ψ(TRI4000+SLP300) | 152.43 | 1.93 | 0.0480 | 0.3810 | 5 | 141.36 |
| ψ(TRI4000+SLP30) | 175.30 | 12.69 | 0.0013 | 0.0018 | 5 | 164.21 | ψ(TRI1000+SLP4000) | 152.44 | 1.94 | 0.0477 | 0.3791 | 5 | 141.37 |
| ψ(TRI2000+SLP2000) | 176.53 | 13.92 | 0.0007 | 0.0009 | 5 | 165.44 | ψ(TRI300+SLP2000) | 152.56 | 2.06 | 0.0449 | 0.3570 | 5 | 141.49 |
| ψ(TRI300+SLP4000) | 177.22 | 14.61 | 0.0005 | 0.0007 | 5 | 166.13 | ψ(TRI1000+SLP30) | 152.61 | 2.11 | 0.0438 | 0.3482 | 5 | 141.54 |
| ψ(TRI30+SLP300) | 178.57 | 15.96 | 0.0002 | 0.0003 | 5 | 167.48 | ψ(TRI2000+SLP300) | 153.23 | 2.73 | 0.0322 | 0.2554 | 5 | 142.16 |
| ψ(TRI2000+SLP4000) | 178.59 | 15.98 | 0.0002 | 0.0003 | 5 | 167.50 | ψ(TRI2000+SLP30) | 153.24 | 2.74 | 0.0320 | 0.2541 | 5 | 142.17 |
| ψ(TRI30+SLP2000) | 178.86 | 16.25 | 0.0002 | 0.0003 | 5 | 167.77 | ψ(TRI300+SLP30) | 153.27 | 2.77 | 0.0315 | 0.2503 | 5 | 142.20 |
| ψ(TRI2000+SLP30) | 178.92 | 16.31 | 0.0002 | 0.0003 | 5 | 167.83 | |||||||
| ψ(TRI300+SLP30) | 179.52 | 16.91 | 0.0002 | 0.0002 | 5 | 168.43 | |||||||
| ψ(TRI30+SLP4000) | 184.43 | 21.82 | 0.0000 | 0.0000 | 5 | 173.34 | |||||||
| ψ(.) | 185.15 | 22.54 | 0.0000 | 0.0000 | 3 | 178.73 |
Note: Akaike's information criterion corrected for small sample sizes (AICc), relative difference in AICc values compared with the top ranked model (ΔAICc), model weight (AICcw), Model likelihood, number of parameters (K), and -2log-likelihood (-2 log LL). Detection covariate was camera trap deployment team (TEAM). Site covariates were: elevation with multi-scale threshold values of 30m, 300 m, 1000 m, 2000 m, 4000 m (ELE30, ELE300, ELE1000, ELE2000, ELE4000), slope with multi-scale threshold values of 30 m, 300 m, 1000 m, 2000 m, 4000 m (SLP30, SLP300, SLP1000, SLP2000, SLP4000), terrain ruggedness index with multi-scale threshold values of 30 m, 300 m, 1000 m, 2000 m, 4000 m (TRI30, TRI300, TRI1000, TRI2000,TRI4000). |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
Snow Leopard China. 2019. Status of snow leopard survey and conservation, China 2018. https://max.book118.com/html/2020/0308/5312013103002230.shtm. in Chinese)
|
| [66] |
Snow Leopard Working Secretariat. 2013. Global snow leopard and ecosystem protection program. Bishkek, Kyrgyz Republic: Snow Leopard Working Secretariat.
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}