
Journal of Resources and Ecology >
Characteristics and Carbon Storage of a Typical Mangrove Island Ecosystem in Beibu Gulf, South China Sea
|
WU Bin, E-mail: wubin@nnnu.edu.cn |
Received date: 2020-11-24
Accepted date: 2021-08-30
Online published: 2022-04-18
Supported by
The Guangxi Science and Technology Base and Talent Project(GuikeAD19245041)
The Guangxi Science and Technology Base and Talent Project(2019AC20088 ))
The Guangxi Key Research and Development Program(AA18118038)
The Project to Improve the Basic Research Ability of Young and Middle-Aged Teachers in Guangxi Universities(2020KY09021)
The Project to Improve the Basic Research Ability of Young and Middle-Aged Teachers in Guangxi Universities(2019KY0426)
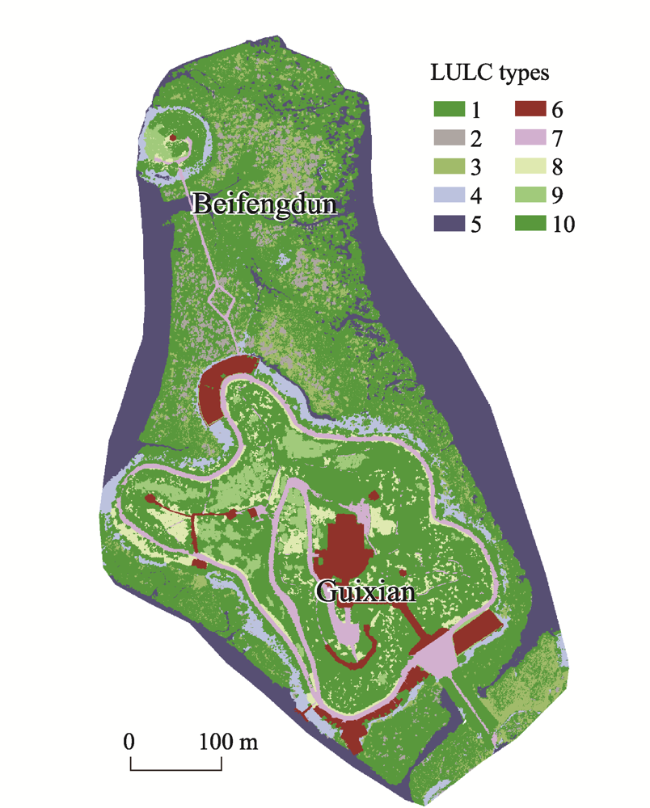
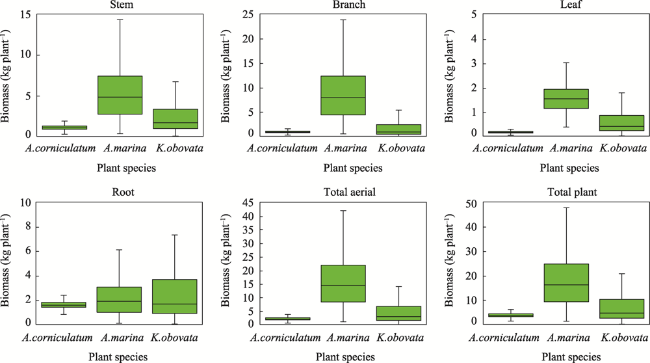
By studying the structural characteristics and carbon storage of the mangrove island ecosystem in the Beibu Gulf, this study provides a scientific basis for mangrove ecological compensation in the coastal areas of Guangxi, South China Sea. On the basis of the unmanned aerial vehicle remote sensing images and a sample plot survey, the object-oriented multi-scale segmentation algorithm is used to extract the mangrove community type information, and one-way analysis of variance is conducted to analyse the structural characteristics of the mangrove community. The carbon storage and carbon density of different mangrove ecosystems were obtained based on the allometric growth equation of mangrove plants. The analysis yielded four main results. (1) The island group covers about 27.10 ha, 41.32% (11.20 ha) of which represents mangrove areas. The mangrove forest is widely distributed in the tidal flats around the islands. (2) The main mangrove types were Aegiceras corniculatum, Kandelia obovata + Aegiceras corniculatum, Avicennia marina + Aegiceras corniculatum and Avicennia marina communities. (3) Amongst the mangrove plants, Avicennia marina had the highest biomass (18.52 kg plant-1), followed by Kandelia obovata (7.84 kg plant-1) and Aegiceras corniculatum (3.85 kg plant-1). (4) The mangrove carbon density difference was significant. Kandelia obovata had the highest carbon density (148.03 t ha-1), followed by Avicennia marina (104.79 t ha-1) and Aegiceras corniculatum (99.24 t ha-1). The carbon storage of the mangrove island ecosystem was 1194.70 t, which was higher than in other areas with the same latitude. The carbon sequestration capacity of the mangrove was relatively strong.
Key words: mangroves; island communities; structural characteristics; biomass; carbon storage
WU Bin , ZHANG Wenzhu , TIAN Yichao , LIANG Mingzhong , XU Jun , GU Guanhai . Characteristics and Carbon Storage of a Typical Mangrove Island Ecosystem in Beibu Gulf, South China Sea[J]. Journal of Resources and Ecology, 2022 , 13(3) : 458 -465 . DOI: 10.5814/j.issn.1674-764x.2022.03.010
Fig. 1 Status of the Guixian and Beifengdun island groupNote: LULC: Land use and land cover. 1. Aegiceras corniculatum; 2. Avicennia marina; 3. Kandelia obovate; 4. Tidal flat; 5. Sea; 6. Construction land; 7. Road; 8. Grassland; 9. Shrub; 10. Forest. |
Table 1 Allometric growth equations of the mangrove plants |
| Plant species | Plant component | Allometric growth equation |
|---|---|---|
| A. corniculatum | Stem | $\ln SB=1.198\text{+}0.464\ln (DB{{H}^{2}}\times HT)$ |
| Branch | $\ln BB=1.110\text{+}0.463\ln (DB{{H}^{2}}\times HT)$ | |
| Leaf | $\ln LB=0.393\text{+}0.475\ln (DB{{H}^{2}}\times HT)$ | |
| Root | $\ln RB=0.967\text{+}0.303\ln (DB{{H}^{2}}\times HT)$ | |
| Total aerial biomass | $\ln TB=1.496\text{+}0.465\ln (DB{{H}^{2}}\times HT)$ | |
| A. marina | Stem | $\ln SB=1.643\text{+}0.544\ln (DB{{H}^{2}}\times HT)$ |
| Branch | $\ln BB=1.897\text{+}0.567\ln (DB{{H}^{2}}\times HT)$ | |
| Leaf | $\ln LB=0.690\text{+}0.287\ln (DB{{H}^{2}}\times HT)$ | |
| Root | $\ln RB=1.361\text{+}0.615\ln (DB{{H}^{2}}\times HT)$ | |
| Total aerial biomass | $\ln TB=2.092\text{+}0.529\ln (DB{{H}^{2}}\times HT)$ | |
| K. obovata | Stem | $\ln SB=2.162\text{+}0.869\ln (DB{{H}^{2}}\times HT)$ |
| Branch | $\ln BB=2.741\text{+}1.253\ln (DB{{H}^{2}}\times HT)$ | |
| Leaf | $\ln LB=1.706\text{+}0.943\ln (DB{{H}^{2}}\times HT)$ | |
| Root | $\ln RB=2.433\text{+}0.990\ln (DB{{H}^{2}}\times HT)$ | |
| Total aerial biomass | $\ln TB=2.814\text{+}1.053\ln (DB{{H}^{2}}\times HT)$ |
Table 2 Mangrove growth characteristics in the plot |
| Plant species | Plant number | Plant height (m) | Breast-height diameter (cm) | Branch count (number) | Crown (m) | Coverage (%) |
|---|---|---|---|---|---|---|
| A. corniculatum | 5527 | 1.97±0.32c | 4.14±1.50c | 5.67±3.96a | 1.06±0.53c | 0.98±3.03a |
| A. marina | 582 | 2.93±0.80a | 8.52±4.56a | 1.67±1.61c | 2.28±1.48b | 0.62±0.32b |
| K. obovata | 839 | 2.59±0.67b | 5.34±2.36b | 3.07±3.10b | 1.29±0.63a | 0.64±0.39ab |
Note: The data are expressed as mean ± standard error; a, b and c indicate significant differences (P<0.05) after the Tukey's honestly significant difference test amongst the different tree species through one-way ANOVA. |
Table 3 Dominant species in the mangrove community |
| Plant species | Relative abundance | Relative significance | Relative frequency | Importance value |
|---|---|---|---|---|
| A. corniculatum | 79.55 | 62.26 | 35.71 | 59.18 |
| A. marina | 12.08 | 11.22 | 33.33 | 18.88 |
| K. obovata | 8.38 | 26.51 | 30.95 | 21.95 |
Table 4 Landscape types in the Guixian and Beifengdun island group |
| Landscape type | Map spots (number) | Perimeter (m) | Area (m2) | Proportion (%) |
|---|---|---|---|---|
| A. corniculatum | 2262 | 56264.87 | 87467.65 | 32.27 |
| A. marina | 1648 | 13714.92 | 8393.98 | 3.10 |
| K. obovate | 5660 | 34476.48 | 16109.60 | 5.94 |
| Tidal flat | 501 | 8162.96 | 10780.81 | 3.98 |
| Sea | 768 | 12808.30 | 47257.78 | 17.44 |
| Construction land | 56 | 2855.68 | 11872.33 | 4.38 |
| Road | 232 | 7073.00 | 13798.78 | 5.09 |
| Grassland | 1201 | 12415.44 | 9912.02 | 3.66 |
| Shrub | 133 | 4510.75 | 8419.65 | 3.11 |
| Forest | 606 | 21762.67 | 57000.52 | 21.03 |
| Total | 13067 | 174045.07 | 271013.12 | 100.00 |
Fig. 2 Biomasses of the mangrove communities (kg plant-1) |
Table 5 Biomasses of mangrove plants (Unit: t ha-1) |
| Plant species | Stem | Branch | Leaf | Root | Total aerial | Total plant |
|---|---|---|---|---|---|---|
| A. corniculatum | 266.57 | 218.89 | 39.29 | 386.12 | 526.49 | 910.87 |
| A. marina | 323.66 | 537.52 | 89.94 | 133.35 | 957.66 | 1084.47 |
| K. obovata | 496.00 | 385.75 | 126.79 | 555.18 | 1027.46 | 1563.72 |
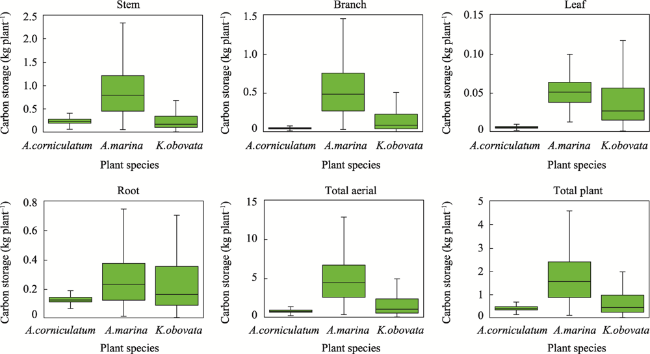
Fig. 3 Carbon storage levels of the mangrove community (kg plant-1) |
Table 6 Carbon density of mangrove plants |
| Plant species | Stem (t ha-1) | Proportion for stem (%) | Branch (t ha-1) | Proportion for branch (%) | Leaf (t ha-1) | Proportion for leaf (%) | Root (t ha-1) | Proportion for root (%) |
|---|---|---|---|---|---|---|---|---|
| A. corniculatum | 57.31 | 57.75 | 10.51 | 10.59 | 1.30 | 1.31 | 30.12 | 30.35 |
| A. marina | 52.76 | 50.35 | 32.79 | 31.29 | 2.97 | 2.83 | 16.27 | 15.53 |
| K. obovata | 50.10 | 33.84 | 36.26 | 24.50 | 8.37 | 5.65 | 53.30 | 36.01 |
Table 7 Carbon storage of mangrove plants |
| Plant species | Area (ha) | Carbon density (t ha-1) | Carbon storage (t) | Proportion (%) |
|---|---|---|---|---|
| A. corniculatum | 8.75 | 99.24 | 868.35 | 72.68 |
| A. marina | 0.84 | 104.79 | 88.02 | 7.37 |
| K. obovata | 1.61 | 148.03 | 238.33 | 19.95 |
| Total | 11.20 | 106.67 | 1194.70 | 100.00 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
FAO. 2008. The world's mangroves 1980-2005. FAO Forestry Paper, (4): 703-704.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}