
Journal of Resources and Ecology >
Spatial Patterns of Soil Organic Matter, Nitrogen, Phosphorus and Potassium in a Subtropical Forest and Its Implication for Forest Management
|
HAN Lili, E-mail: Lhan0325@163.com |
Received date: 2020-10-20
Accepted date: 2021-09-08
Online published: 2022-04-18
Supported by
The National Key Research & Development Program of China(2016YFD060020501)
Total nitrogen (TN), total phosphorus (TP), total potassium (TK), and soil organic matter (OM) can significantly affect forest growth. However, these soil properties are spatially heterogeneously distributed, complicating the prescription of forest management strategies. Thus, it is imperative to obtain an in-depth understanding of the spatial distribution of soil properties. In this study, soils were sampled at 181 locations in the Tropical Forest Research Center in the southwestern Guangxi Zhuang Autonomous Region in southern China. We investigated the spatial variability of soil OM, TN, TP, and TK using geostatistical analysis. The nugget to sill ratio indicated a strong spatial dependence of soil TN and a moderate spatial dependence of soil OM, TP, and TK, suggesting that TN was primarily controlled by intrinsic factors (e.g., soil texture, parent material, vegetation type, and topography), whereas soil OM, TP, and TK were controlled by intrinsic and extrinsic factors (e.g., cultivation practices, fertilization, and planting systems). Based on the spatial variability determined by the geostatistical analysis, we performed ordinary kriging to create thematic maps of soil TN, TP, TK, and OM. Model validation indicated that the thematic maps were reliable to inform forest management.
HAN Lili , LU Yuanchang , MA Wu , MENG Jinghui . Spatial Patterns of Soil Organic Matter, Nitrogen, Phosphorus and Potassium in a Subtropical Forest and Its Implication for Forest Management[J]. Journal of Resources and Ecology, 2022 , 13(3) : 417 -427 . DOI: 10.5814/j.issn.1674-764x.2022.03.007
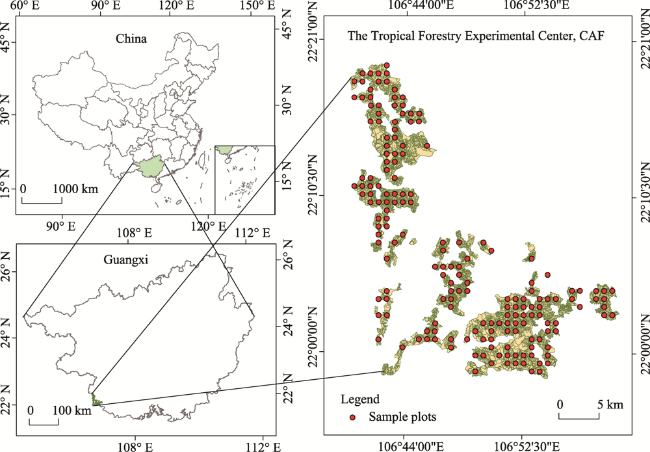
Fig. 1 Distribution of the 181 systematic sample plots in the study area |
Table 1 Descriptive statistics of soil TN, TP, TK, and OM in the 0-60 cm soil horizon. (Unit: g kg-1) |
| Nutrient | Mean | Min | Max | Median | SD | CV (%) | P-value of S-W |
|---|---|---|---|---|---|---|---|
| OM | 21.66 | 6.61 | 58.25 | 20.05 | 9.23 | 42.62 | <0.0001 |
| TN | 1.10 | 0.35 | 2.50 | 1.09 | 0.36 | 32.75 | <0.0001 |
| TP | 0.26 | 0.10 | 0.74 | 0.24 | 0.10 | 39.04 | <0.0001 |
| TK | 9.78 | 0.61 | 38.03 | 9.58 | 7.27 | 74.38 | <0.0001 |
Note: Min: Minimum value; Max: Maximum value; SD: Standard deviation; CV: Coefficient of variance; S-W test: Shapiro-Wilk test; OM: Organic matter density; TN: Total nitrogen density; TP: Total phosphorus density; TK: Total potassium density. |
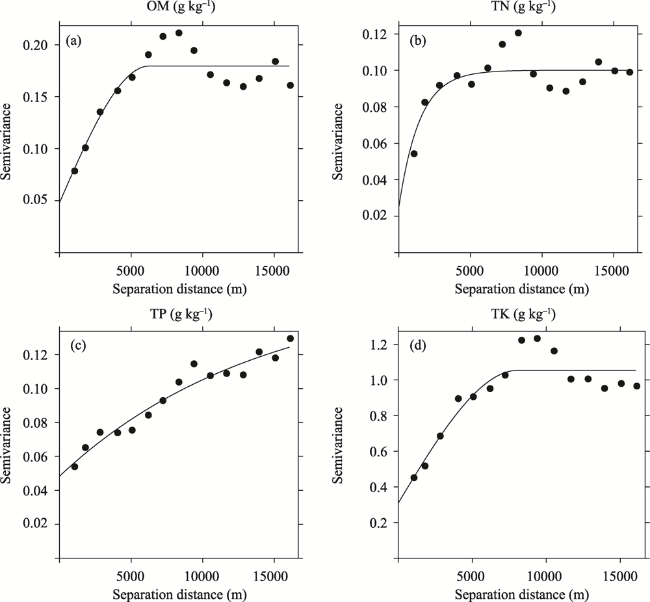
Fig. 2 Experimental semivariograms of soil OM (a), TN (b), TP (c) and TK (d). |
Table 2 Models and fitted parameters of the semivariograms of soil OM, TN, TP, and TK. (Unit: g kg-1) |
| Nutrient | Models | Nugget (C0) | Sill (C0+C) | Nugget/Sill | Range (A0, m) | Determination coefficient |
|---|---|---|---|---|---|---|
| OM | Spherical | 0.0480 | 0.1794 | 0.27 | 6250 | 0.815 |
| TN | Exponential | 0.0250 | 0.1001 | 0.25 | 4347 | 0.655 |
| TP | Exponential | 0.0483 | 0.1598 | 0.30 | 14015 | 0.948 |
| TK | Spherical | 0.3098 | 1.0546 | 0.29 | 8114 | 0.848 |
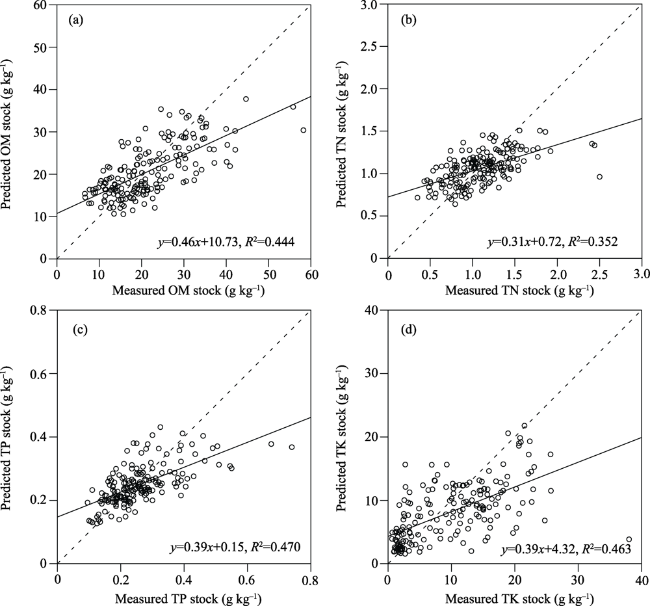
Fig. 3 Cross-validation of kriging interpolation for OM (a), TN (b), TP (c) and TK(d) (dashed line denotes the 1:1 line). |
Table 3 The lack-of-fit statistics of the semivariogram model of soil OM, TN, TP, and TK. (Unit: g kg-1) |
| Nutrient | AME | ME | RMSE |
|---|---|---|---|
| OM | 0.2615 | 0.0017 | 0.3185 |
| TN | 0.2132 | 0.0028 | 0.2704 |
| TP | 0.2082 | 0.0002 | 0.2626 |
| TK | 0.5645 | 0.0127 | 0.7192 |
Note: AME: Absolute mean error; ME: Mean error; RMSE: Root mean square error. |
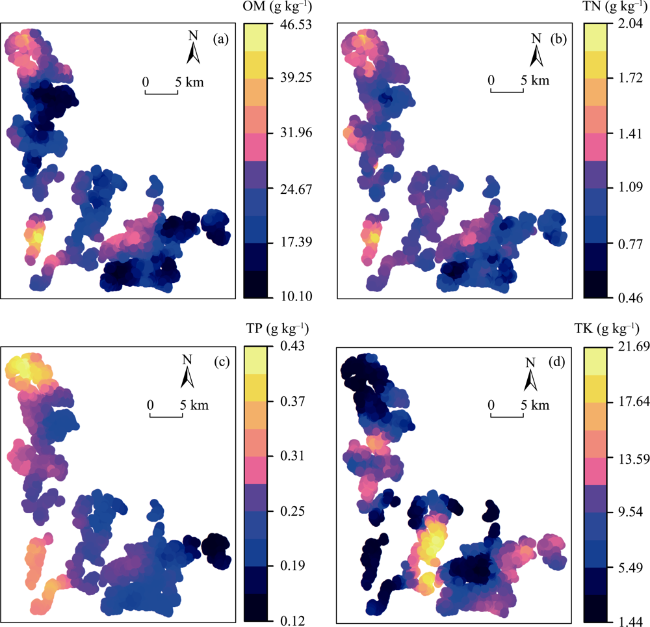
Fig. 4 The thematic maps of soil OM (a), TN (b), TP (c) and TK (d) in the TFEC obtained by ordinary kriging. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
State Forestry Adiministration. 1999. Forestry standard of People's Republic of China-Methods of forest soil analysis. Beijing, China: Chinese Standard Press. (in Chinese)
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}