
Journal of Resources and Ecology >
Seed and Fruiting Phenology Plasticity and Offspring Seed Germination Rate in Two Asteraceae Herbs Growing in Karst Soils with Varying Thickness and Water Availability
|
LIU Junting, E-mail: 1109547757@qq.com |
Received date: 2021-03-24
Accepted date: 2021-10-19
Online published: 2022-03-09
Supported by
The Chongqing Natural Science Foundation(cstc2020jcyj-msxmX0244)
The Special Funds for Basic Scientific Research in Central Universities(XDJK2020B037)
The National Natural Science Foundation of China(31500399)
Shallow soil with low water availability is the key limiting factor for plant growth and reproduction in vulnerable karst regions. Annual herbs are pioneers adapted to these areas; however, little is known about the responses of their seeds and infructescence, and the germination of their offspring to these limited water and soil resources. In this study, we investigated how the seed and fruiting phenology plasticity and offspring seed germination rates of two annual Asteraceae herbs (Xanthium sibiricum and Bidens pilosa) respond to the harsh karst soil environment, by assessing the seed number, seed biomass and offspring seed germination rate. X. sibiricum and B. pilosa were grown under three soil thicknesses and three water availability levels in a full two-way randomized block design. The key results were as follows: (1) The number and biomass of progenies (infructescence and seeds) of X. sibiricum decreased with the decline of soil thickness and/or water availability (P <0.05). The infructescence and seed biomass of B. pilosa increased with the decline of water availability. (2) Seed quantity and seed biomass of X. sibiricum showed no correlation after their parents experienced resource reductions. A significant positive relationship between seed number and seed biomass was observed in B. pilosa (P <0.05). (3) The offspring seed germination rate of X. sibiricum did not change with the decrease of soil thickness under three levels of water treatment. However, the offspring seed germination rate of B. pilosa decreased significantly with the decrease of soil thickness under the control water level (P<0.05). The results show that X. sibiricum tends to improve its competitiveness by ensuring the quantity and quality of offspring in order to adapt to the shallow karst soils and dry karst habitats. In contrast, B. pilosa adapts to the unfavorable karst habitats by a risk-sharing strategy. B. pilosa produces more and bigger seeds to in an attempt to expand its survival range and escape from the unfavorable living environment, but this results in a lower seed number and germination rate of its progeny under the karst soil resource reduction.
Key words: karst drought; shallow soil; seed number; parental environment; seed biomass
LIU Junting , LI Suhui , SONG Haiyan , LEI Ying , CHEN Jinyi , WANG Jiamin , GUO Xuman , LIU Jinchun . Seed and Fruiting Phenology Plasticity and Offspring Seed Germination Rate in Two Asteraceae Herbs Growing in Karst Soils with Varying Thickness and Water Availability[J]. Journal of Resources and Ecology, 2022 , 13(2) : 319 -327 . DOI: 10.5814/j.issn.1674-764x.2022.02.014
Table 1 Water design during the test |
| Months | WCK | WM | WL |
|---|---|---|---|
| Apr.-Jun. | 130 ml (3 d) -1 | 65 ml (3 d) -1 | 43 ml (3 d) -1 |
| Jul.-Sept. | 125 ml (3 d) -1 | 63 ml (3 d) -1 | 38 ml (3 d) -1 |
| Oct.-Dec. | 56 ml (3 d) -1 | 28 ml (3 d) -1 | 19 ml (3 d) -1 |
Note: WCK, WM and WL: high (no reduction in water), moderate (50% reduction in water) and low (70% reduction in water) water availability, respectively. |
Table 2 Relative soil water content under different treatments |
| Water application | ||||||
|---|---|---|---|---|---|---|
| The thickness of the soil | WCK | WM | WL | |||
| Soil moisture content (%) | Relative water content | Soil moisture content (%) | Relative water content | Soil moisture content (%) | Relative water content | |
| SCK | 19.16±0.71Aa | 48%FC | 14.13±0.32Ba | 36%FC | 10.81±0.33Ca | 27%FC |
| SM | 20.55±0.53Aa | 51%FC | 12.86±0.46Ba | 32%FC | 9.36±0.70Ca | 24%FC |
| SL | 21.05±0.57Aa | 52%FC | 12.60±0.71Ba | 32%FC | 7.47±0.38Cb | 19%FC |
Note: WCK, WM and WL: high (no reduction in water), moderate (50% reduction in water) and low (70% reduction in water) water availability, respectively. SCK, SM and SL: deep (70 cm), moderate (40 cm) and shallow (10 cm) thickness of the soil, respectively. Lowercase letters indicate significant differences between different thicknesses of the soil at the 0.05 level, and uppercase letters indicate significant differences between different water availability levels at the 0.05 level. FC: field water holding capacity. |
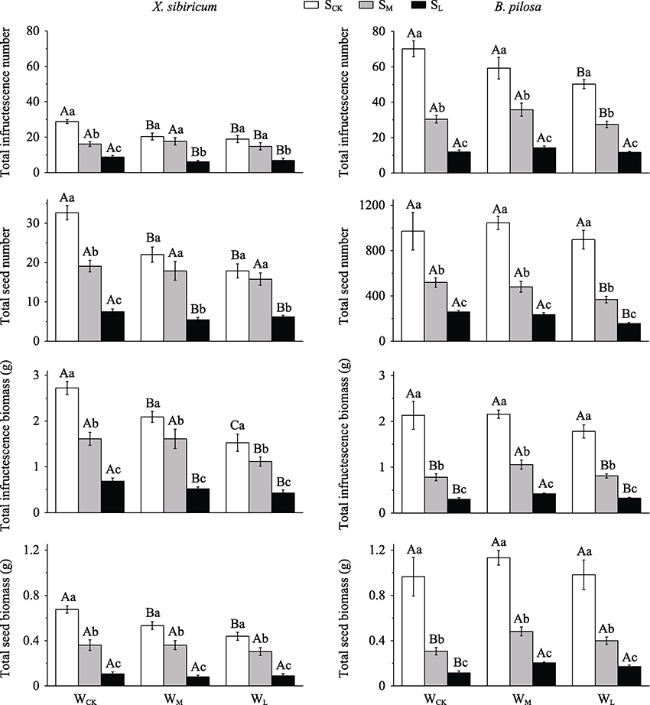
Fig. 1 Effects of different thicknesses of the soil and water availability on infructescence (and seed) number and biomass of X. sibiricum and B. pilosa (M±SE)Notes: WCK, WM and WL: high (no reduction in water), moderate (50% reduction in water) and low (70% reduction in water) water availability, respectively. SCK, SM and SL: deep (70 cm), moderate (40 cm) and shallow (10 cm) thickness of the soil, respectively. Different lowercase letters indicate significant differences between different thicknesses of the soil under the same water availability (P < 0.05). Different uppercase letters indicate significant differences between different water availability levels under the same thickness of the soil. |
Table 3 Results of Two-way ANOVA on the infructescence and seed number and biomass of X. sibiricum and B. pilosa |
| Species | Treatment | F-value | |||
|---|---|---|---|---|---|
| Infructescence number | Seed number | Infructescence biomass | Seed biomass | ||
| X. sibiricum | Water | 5.36** | 11.72** | 15.26** | 6.29** |
| Soil | 58.43** | 78.79** | 85.97** | 118.18** | |
| Water × Soil | 3.18* | 5.11** | 3.26* | 2.95* | |
| B. pilosa | Water | 7.55** | 4.56* | 4.83* | 4.97* |
| Soil | 216.49** | 142.38** | 212.94** | 162.22** | |
| Water × Soil | 4.10** | 0.34ns | 1.36ns | 0.38ns | |
Note: *, ** indicate significant differences at P < 0.05, P < 0.01, respectively. ns: no significant difference at the 0.05 level. |
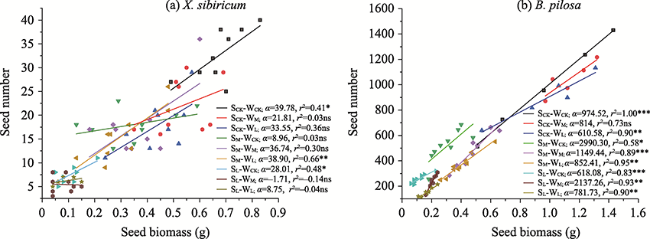
Fig. 2 The relationship between seed number and seed biomass of X. sibiricum (a) and B. pilosa (b)Note: r: Pearson’s correlation coefficient; α: Regression slope. WCK, WM and WL: high (no reduction in water), moderate (50% reduction in water) and low (70% reduction in water) water availability, respectively. SCK, SM and SL: deep (70 cm), moderate (40 cm) and shallow (10 cm) thickness of the soil, respectively; *, ** and *** indicate significant differences at P < 0.05, P < 0.01 and P < 0.001, respectively; ns: no significant difference at the 0.05 level. |
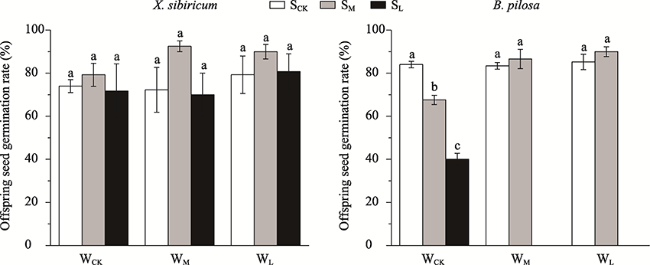
Fig. 3 The effect of different thicknesses of soil and water availability levels on offspring seed germination rates of X. sibiricum and B. pilosaNote: WCK, WM and WL: high (no reduction in water), moderate (50% reduction in water) and low (70% reduction in water) water availability, respectively. SCK, SM and SL: deep (70 cm), moderate (40 cm) and shallow (10 cm) thickness of the soil, respectively. Different lowercase letters indicate significant differences between the different thicknesses of soil under the same water availability level (P < 0.05). |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}