
Journal of Resources and Ecology >
Study on Seed Bank Composition and Seedling Emergence Regularity in Placer Iron Ore Soil
|
HOU Yurong, E-mail: houyurong0994@126.com |
Received date: 2021-03-15
Accepted date: 2021-08-02
Online published: 2022-03-09
Supported by
The Project of Xinjiang Uygur Autonomous Region(2020D01A38)
The mining of placer iron ore greatly influences the surrounding desert grassland. In Agdala Town, Qinghe County, Xinjiang, the soil seed bank is severely damaged, and the utilization and productivity of desert grasslands are almost zero, which seriously affects the safety of the ecological environment and the development of local animal husbandry. It is very important to supplement soil seed banks to enhance the species composition of desert grasslands after ecological restoration. In this study, the effects of the seed bank, species composition, and seed burial depth on the seedling emergence rate at placer iron ore site before and after artificial seed bank replenishment were compared using the germination tray method. The original soil seed bank in the study area contained only four species, which were all annual plants. The dominant species were Salsola ruthenica and Corispermum orientale, and the soil seed bank density was very small. The emergence regularity of the soil seed bank was closely related to water content, and the emergence of annual vegetation was explosive. Seed burial depth affected the emergence rate of perennial grass, and we determined that a burial depth of 0-5 cm was the most effective for emergence. This indicates that seedling emergence of artificially supplemented soil seed bank can be completed within 7 days if the sowing depth is appropriate; sowing depths greater than 5 cm affect seed germination. These findings provide a basis for selecting species to populate large areas.
Key words: seedling emergence; placer iron ore; soil seed band; species composition
HOU Yurong , KE Mei , WEI Peng , LAN Jiyong , LI Chao , KANG Shuai , KAYRAT Aldeyarhan , WANG Lu . Study on Seed Bank Composition and Seedling Emergence Regularity in Placer Iron Ore Soil[J]. Journal of Resources and Ecology, 2022 , 13(2) : 312 -318 . DOI: 10.5814/j.issn.1674-764x.2022.02.013
Table 1 Primitive soil seed bank composition of the top 15 cm soil layer |
| Soil depth | Plant name | ||||
|---|---|---|---|---|---|
| Salsola ruthenica | Corispermum orientale | Atriplex cana | Petrosimonia sibirica | Bassia dasyphylla | |
| 0-5 cm | 51 ± 0.48Aa | 29 ± 0.41Bb | 21 ± 0.41Cc | 3 ± 0.41Dd | 0 ± 0.00Ee |
| 5-10 cm | 55 ± 0.88Aa | 25 ± 0.58Bb | 7 ± 0.33Cc | 3 ± 0.00Dd | 1 ± 0.58De |
| 10-15 cm | 48 ± 0.58Aa | 36 ± 0.33Bb | 7 ± 0.33Cc | 2 ± 0.67Dd | 2 ± 0.58Dd |
Note: Lowercase letters indicate significant difference at 5% level and uppercase letters indicate significant difference at 1% level. |
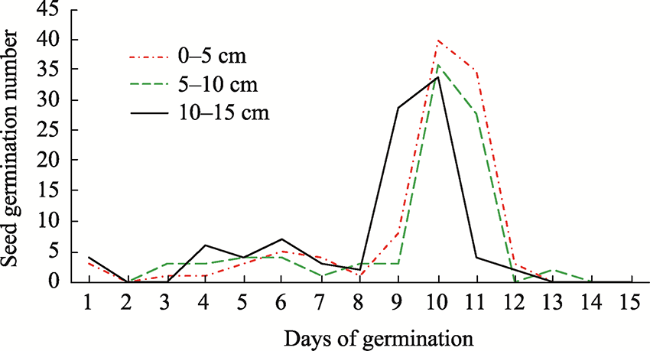
Fig. 1 Germination number of seeds in each soil layer |
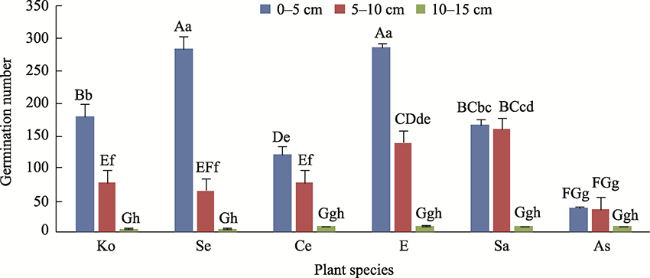
Fig. 2 Plant composition of artificially reseeded soil at sowing depths of 0-5 cm, 5-10 cm, and 10-15 cm.Note: Ko: Kochia prostrata; Se: Seriphidium sp.; Ce: Ceratoides latens; E: Elytrigia repens; Sa: Salsola collina; As: Astragalus membranaceus. |
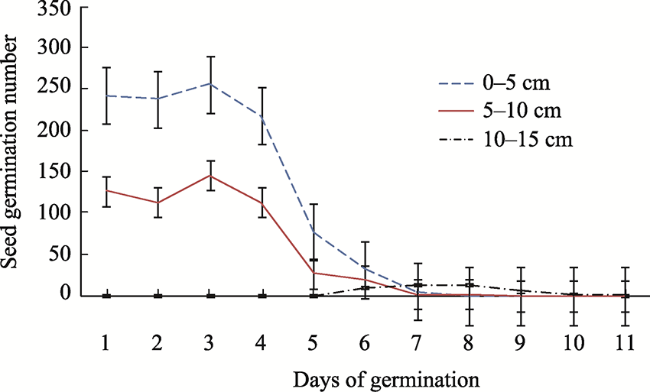
Fig. 3 Total seedling emergence of the 0-15 cm soil layerNote: Plant names are defined in Fig. 2. |
Fig. 4 Emergence frequency of artificially replenished soil seed bank at sowing depths of 0-5 cm, 5-10 cm, and 10- 15 cm. |
We thank Lin Hongkai for choosing the location of the research area and providing useful suggestions. This work was partially funded by the Key Technologies of Ecological Restoration of Degraded Grasslands as a Part of the “Study on the Key Technologies of Ecological Protection and Restoration of the Community of Lakes and Grasses in the Mountains, Rivers, Forests, and Fields of the Drainage Area” and Effects of Different Ecological Restoration Modes on Soil Nutrients and Microorganisms in Junggar Desert.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}