
Journal of Resources and Ecology >
Predicting the Impact of Climate Change on Vulnerable Species in Gandaki River Basin, Central Himalayas
|
Raju RAI, E-mail: rairaju@igsnrr.ac.cn |
Received date: 2021-05-28
Accepted date: 2021-10-30
Online published: 2022-03-09
Supported by
The Second Tibetan Plateau Scientific Expedition and Research(2019QZKK0603)
The Strategic Priority Research Program of Chinese Academy of Sciences(XDA20040201)
The National Natural Science Foundation of China(41761144081)
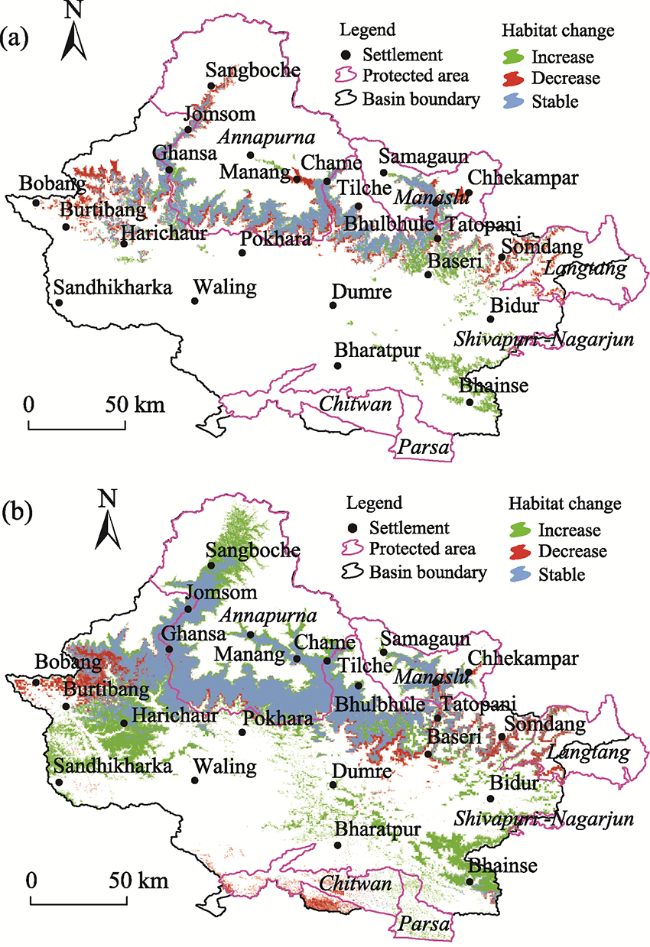
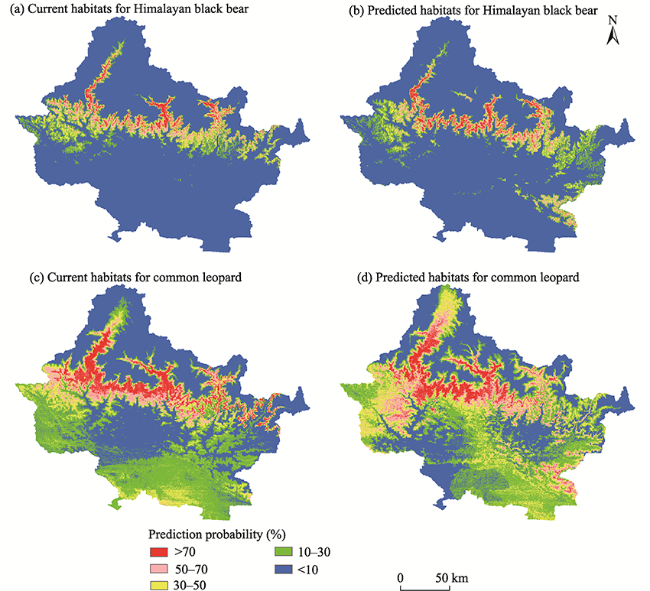
Gandaki River Basin (GRB) is an important part of the central Himalayan region, which provides habitat for numerous wild species. However, climatic changes are making the habitat in this basin more vulnerable. This paper aims to assess the potential impacts of climate change on the spatial distributions of habitat changes for two vulnerable species, Himalayan black bear (Ursus thibetanus laniger) and common leopard (Panthera pardus fusca), using the maximum entropy (MaxEnt) species distribution model. Species occurrence locations were used along with several bioclimatic and topographic variables (elevation, slope and aspect) to fit the model and predict the potential distributions (current and future) of the species. The results show that the highly suitable area of Himalayan black bear within the GRB currently encompasses around 1642 km2 (5.01% area of the basin), which is predicted to increase by 51 km2 in the future (2050). Similarly, the habitat of common leopard is estimated as 3999 km2 (12.19% of the GRB area), which is likely to increase to 4806 km2 in 2050. Spatially, the habitat of Himalayan black bear is predicted to increase in the eastern part (Baseri, Tatopani and north from Bhainse) and to decrease in the eastern (Somdang, Chhekampar), western (Burtibang and Bobang) and northern (Sangboche, Manang, Chhekampar) parts of the study area. Similarly, the habitat of common leopard is projected to decrease particularly in the eastern, western and southern parts of the basin, although it is estimated to be extended in the southeastern (Bhainse), western (Harichaur and northern Sandhikhark) and north-western (Sangboche) parts of the basin. To determine the habitat impact, the environmental variables such as elevation, Bio 15 (precipitation seasonality) and Bio 16 (precipitation of wettest quarter) highly contribute to habitat change of Himalayan black bear; while Bio 13 (precipitation of wettest month) and Bio 15 are the main contributors for common leopard. Overall, this study predicted that the suitable habitat areas of both species are likely to be impacted by climate change at different altitudes in the future, and these are the areas that need more attention in order to protect these species.
Raju RAI , ZHANG Yili , LIU Linshan , Paras Bikram SINGH , Basanta PAUDEL , Bipin Kumar ACHARYA , Narendra Raj KHANAL . Predicting the Impact of Climate Change on Vulnerable Species in Gandaki River Basin, Central Himalayas[J]. Journal of Resources and Ecology, 2022 , 13(2) : 173 -185 . DOI: 10.5814/j.issn.1674-764x.2022.02.001
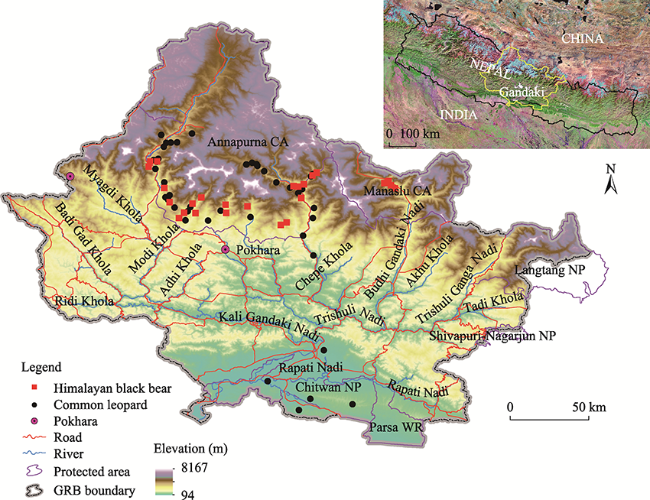
Fig. 1 Location of GRB and species occurrencesNote: CA-Conservation Area; NP-National Park; WR-Wildlife Reserve. |
Table 1 Environmental variables used in this study and their descriptions |
| Variable | Description | Himalayan black bear | Common leopard | |
|---|---|---|---|---|
| Climate | Bio 1 | Annual mean temperature | √ | × |
| Bio 2 | Mean diurnal range | √ | × | |
| Bio 3 | Isothermality | √ | × | |
| Bio 4 | Temperature seasonality | × | × | |
| Bio 5 | Max temperature of warmest month | × | × | |
| Bio 6 | Min temperature of coldest month | √ | √ | |
| Bio 7 | Temperature annual range (Bio 5, 6) | × | × | |
| Bio 8 | Mean temperature of wettest quarter | × | × | |
| Bio 9 | Mean temperature of driest quarter | √ | √ | |
| Bio 10 | Mean temperature of warmest quarter | × | √ | |
| Bio 11 | Mean temperature of coldest quarter | √ | √ | |
| Bio 12 | Annual precipitation | × | √ | |
| Bio 13 | Precipitation of wettest month | √ | √ | |
| Bio 14 | Precipitation of driest month | √ | √ | |
| Bio 15 | Precipitation seasonality | √ | √ | |
| Bio 16 | Precipitation of wettest quarter | √ | √ | |
| Bio 17 | Precipitation of driest quarter | √ | √ | |
| Bio 18 | Precipitation of warmest quarter | √ | × | |
| Bio 19 | Precipitation of coldest quarter | √ | × | |
| Topography | Elevation, slope, aspect | √ | √ | |
Note: √- retained; ×- excluded |
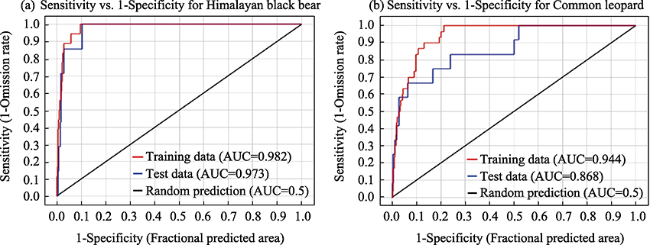
Fig. 2 The AUC of habitat prediction (the training and test data are denoted by red and blue lines, respectively) |
Table 2 Environmental variables and their contributions (%) to the species habitats |
| Variables | Species | |
|---|---|---|
| Himalayan black bear | Common leopard | |
| Bio 1 | 0 | × |
| Bio 2 | 0.5 | × |
| Bio 3 | 0.7 | × |
| Bio 6 | 6.6 | 2.3 |
| Bio 9 | 0 | 4.3 |
| Bio 10 | × | 1.5 |
| Bio 11 | 0 | 0 |
| Bio 12 | × | 1.4 |
| Bio 13 | 0.5 | 39.8 |
| Bio 14 | 0 | 4.6 |
| Bio 15 | 19.1 | 26.8 |
| Bio 16 | 17.3 | 1.3 |
| Bio 17 | 1.3 | 0.2 |
| Bio 18 | 0 | × |
| Bio 19 | 11.5 | × |
| Elevation | 36.8 | 14.3 |
| Slope | 0.1 | 2.6 |
| Aspect | 5.7 | 1.0 |
Note: × - excluded |
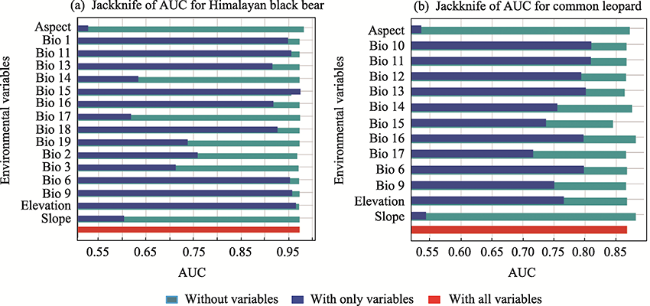
Fig. 3 Jackknife AUC of the different environmental variables |
Fig. 4 Predicting the changes in the habitat suitability within the GRB for the Himalayan black bear (a) and common leopard (b) |
Table 3 Habitat status within the GRB (Unit: km2) |
| Habitat change | Himalayan black bear | Common leopard |
|---|---|---|
| Increase | 1035.05 | 3894.33 |
| Decrease | 1045.86 | 1257.37 |
| Stable | 1985.56 | 4504.81 |
| Net change | 1974.75 | 7141.78 |
Table 4 Current and future predicted habitat in the GRB (Unit: km2) |
| Habitat suitability | Himalayan black bear | Common leopard | ||
|---|---|---|---|---|
| Current | Future | Current | Future | |
| Very high | 616.38 | 692.35 | 1973.62 | 1888.06 |
| High | 1025.76 | 1000.15 | 2024.99 | 2917.51 |
| Medium | 1482.78 | 1565.61 | 2731.43 | 4600.69 |
| Low | 2459.98 | 2814.51 | 7671.67 | 8603.56 |
| Very low | 27222.90 | 26734.50 | 18407.00 | 14797.42 |
Fig. 5 Current and future predicted habitats of the Himalayan black bear (a, b); and common leopard (c, d) |
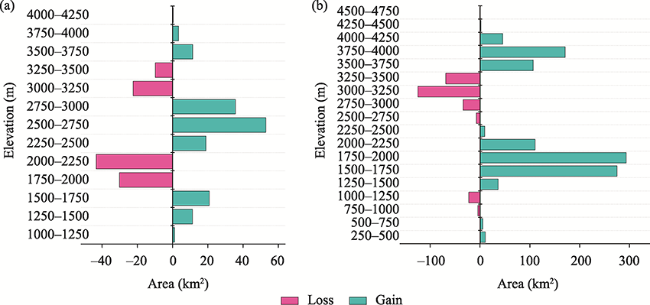
Table 5 Predicting habitat changes along the altitudinal gradient (Unit: km2) |
| Elevation (m) | Species and habitat | Gain and loss | ||||
|---|---|---|---|---|---|---|
| Himalayan black bear (HBB) | Common leopard (CL) | |||||
| Current | Future | Current | Future | HBB | CL | |
| 250-500 | 0.00 | 10.71 | 10.71 | |||
| 500-750 | 0.62 | 6.58 | 5.96 | |||
| 750-1000 | 6.35 | 2.09 | -4.26 | |||
| 1000-1250 | 0.36 | 1.56 | 52.85 | 30.48 | 1.20 | -22.37 |
| 1250-1500 | 9.98 | 21.32 | 165.85 | 202.38 | 11.34 | 36.54 |
| 1500-1750 | 46.89 | 67.72 | 220.20 | 495.32 | 20.82 | 275.12 |
| 1750-2000 | 118.19 | 87.98 | 268.65 | 561.90 | -30.21 | 293.26 |
| 2000—2250 | 210.71 | 167.38 | 348.68 | 459.18 | -43.34 | 110.50 |
| 2250-2500 | 282.01 | 301.02 | 384.52 | 394.12 | 19.01 | 9.59 |
| 2500—2750 | 331.46 | 384.46 | 418.93 | 411.71 | 53.00 | -7.23 |
| 2750-3000 | 295.66 | 331.31 | 483.83 | 449.92 | 35.65 | -33.91 |
| 3000—3250 | 244.07 | 221.63 | 612.11 | 487.54 | -22.43 | -124.57 |
| 3250-3500 | 96.30 | 86.45 | 555.95 | 487.36 | -9.85 | -68.59 |
| 3500-3750 | 6.49 | 17.91 | 341.66 | 448.69 | 11.41 | 107.02 |
| 3750-4000 | 0.00 | 3.43 | 115.23 | 286.02 | 3.43 | 170.79 |
| 4000-4250 | 0.00 | 0.30 | 21.09 | 67.05 | 0.30 | 45.96 |
| 4250-4500 | 1.45 | 3.61 | 2.16 | |||
| 4500-4750 | 0.63 | 0.00 | -0.63 | |||
Fig. 6 Potential changes in the highly suitable (i.e., very high and high probability) habitat of the species at the different elevation ranges for Himalayan black bear (a) and common leopard (b) |
This study was financially supported in part by Chinese Academy of Sciences-The World Academy of Sciences (CAS-TWAS) President’s Fellowship Program for PhD Study. We are thankful to National Trust for Nature Conservation (NTNC), Annapurna Conservation Area Project (ACAP) for providing species data.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
DNPWC. Tiger and prey base monitoring protocol 2017 (Nepal). Department of National Parks and Wildlife Conservation (DNPWC), Ministry of Forests and Soil Conservation, Kathmandu, Nepal. https://dnpwc.gov.np/media/publication/Tiger-and-prey-base-monitoring-protocol.pdf
|
| [19] |
DNPWC. 2018. Protected areas of Nepal. Department of National Parks and Wildlife Conservation (DNPWC), Kathmandu, Nepal.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
IPCC. 2013. Summary for policymakers. In: Stocker T, Qin D, Plattner G K. et al. (eds) Climate change 2013: The physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York, USA: Cambridge University Press.
|
| [34] |
IPCC. 2018. Summary for Policymakers. In: Masson-Delmotte V, Zhai P, Pörtner H O, et al. (eds.). Global warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. Geneva, Switzerland: World Meteorological Organization.
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
MoFE (Ministry of Forests and Environment of Nepal). 2018. Nepal’s Convention on International Trade in Endangered Species (CITES) Wild Fauna and Flora. Kathmandu, Nepal: Government of Nepal, Ministry of Forests and Environment, Department of National Park and Conservation.
|
| [53] |
MoPE (Ministry of Population and Environment of Nepal). 2018. Weather summary of Nepal Year-2014. Kathmandu, Nepal: Government of Nepal, Ministry of Population and Environment, Department of Hydrology and Meteorology.
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}