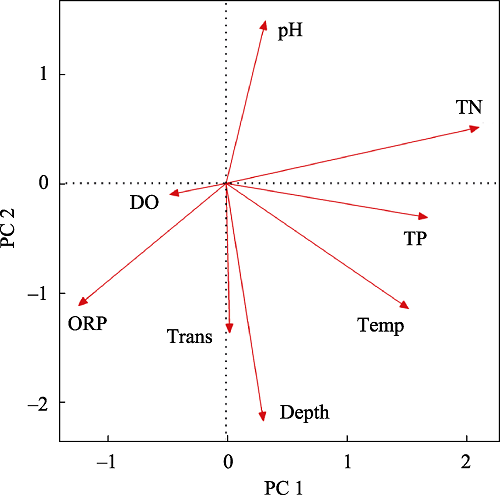
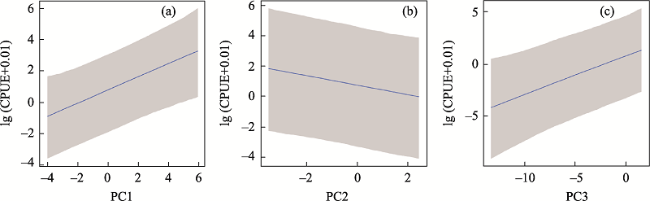
Water column TN and TP were positively correlated with the
P. clarkii CPUE, because as nutrient contents increase, the primary productivity increases correspondingly. On the other hand,
P. clarkii activities such as burrowing and feeding would destroy the growth of large aquatic plants and increase the resuspension of sediments, so an increase in its abundance would aggravate the transition of shallow lakes from clear water dominated by large aquatic plants to turbid water dominated by phytoplankton, thus aggravating the eutrophication of water bodies (Matsuzaki et al.,
2009).
P. clarkii CPUE was negatively related with the DO content, which was consistent with studies on the rivers of Louisiana, USA, the hometown of
P. clarkii, where the individual size and average CPUE of crayfish in a hypoxic environment are both smaller than in a normal dissolved oxygen environment (Bonvillain et al.,
2015). Water ORP determines the forms of water nutrients and plays an important role in nutrient circulation. Its value can affect the oxidation degree of sediment, and thus affect the survival rate of macrobenthos which are also important food resources of
P. clarkii. The results of this study indicated that
P. clarkii CPUE was positively correlated with temperature. A review of
P. clarkii global population temperature tolerance found that its optimal growth temperature range is 21.8-26.7 ℃, and its highest temperature tolerated is 35 ℃ (Westhoff and Rosenberger,
2016).
P. clarkii is a native of tropical regions while the middle-low Yangtze River is a subtropical region, therefore its abundance showed an increasing trend with temperature. This study also found that the
P. clarkii CPUE was positively correlated with water depth, which was consistent with the findings in the Pacific Northwest lakes of the United States (Larson and Olden,
2013), probably because a deep water depth could provide a more diversified habitat for
P. clarkii as refuge. Submerged plant abundance at WDTL was positively related with water transparency (Guo et al.,
2020), and since such plants provide shelter for living benthic animals,
P. clarkii CPUE was also positively related to water transparency. Although
P. clarkii can tolerate a low pH environment through its hard exoskeleton, as the pH decreases, phytoplankton, zooplankton and macrobenthos that are sensitive to water acidification would disappear, resulting in insufficient food resources for
P. clarkii. For example, snails disappear when the pH drops to 5.2 due to an insufficient availability of Ca(HCO
3)
2. A survey of 54 American lakes found that average pH of lakes with
P. clarkii survival was 7.2 (Larson and Olden,
2013).

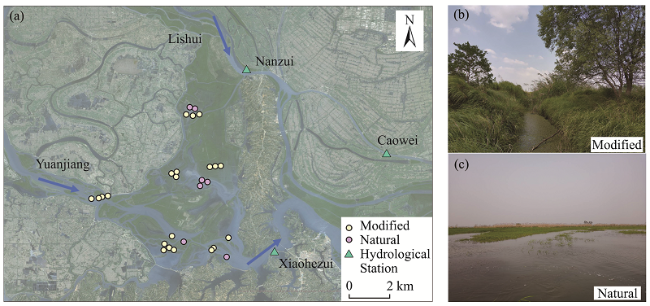
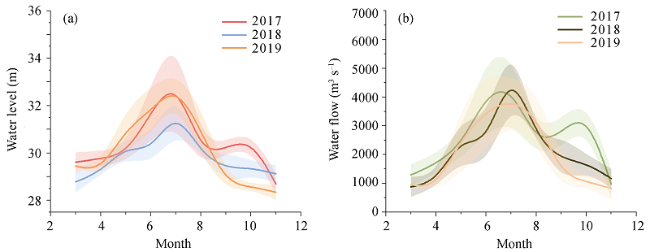
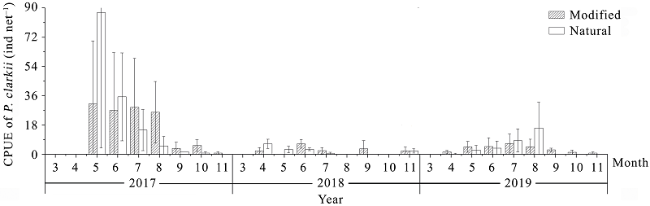
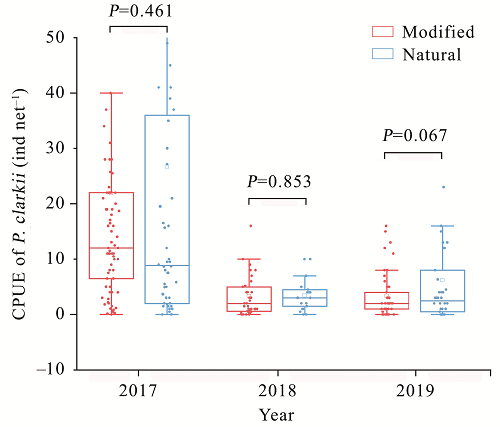
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}