
Journal of Resources and Ecology >
Terrestrial Ecosystem Modeling with IBIS: Progress and Future Vision
Received date: 2021-08-16
Accepted date: 2021-10-14
Online published: 2022-01-08
Supported by
The Key Project of National Natural Science Foundation of China(41930651)
The National Natural Science Foundation of China(41871334)
Dynamic Global Vegetation Models (DGVM) are powerful tools for studying complicated ecosystem processes and global changes. This review article synthesizes the developments and applications of the Integrated Biosphere Simulator (IBIS), a DGVM, over the past two decades. IBIS has been used to evaluate carbon, nitrogen, and water cycling in terrestrial ecosystems, vegetation changes, land-atmosphere interactions, land-aquatic system integration, and climate change impacts. Here we summarize model development work since IBIS v2.5, covering hydrology (evapotranspiration, groundwater, lateral routing), vegetation dynamics (plant functional type, land cover change), plant physiology (phenology, photosynthesis, carbon allocation, growth), biogeochemistry (soil carbon and nitrogen processes, greenhouse gas emissions), impacts of natural disturbances (drought, insect damage, fire) and human induced land use changes, and computational improvements. We also summarize IBIS model applications around the world in evaluating ecosystem productivity, carbon and water budgets, water use efficiency, natural disturbance effects, and impacts of climate change and land use change on the carbon cycle. Based on this review, visions of future cross-scale, cross-landscape and cross-system model development and applications are discussed.
Key words: IBIS; ecological model; productivity; carbon cycle; global change
LIU Jinxun , LU Xuehe , ZHU Qiuan , YUAN Wenping , YUAN Quanzhi , ZHANG Zhen , GUO Qingxi , DEERING Carol . Terrestrial Ecosystem Modeling with IBIS: Progress and Future Vision[J]. Journal of Resources and Ecology, 2022 , 13(1) : 2 -16 . DOI: 10.5814/j.issn.1674-764x.2022.01.001
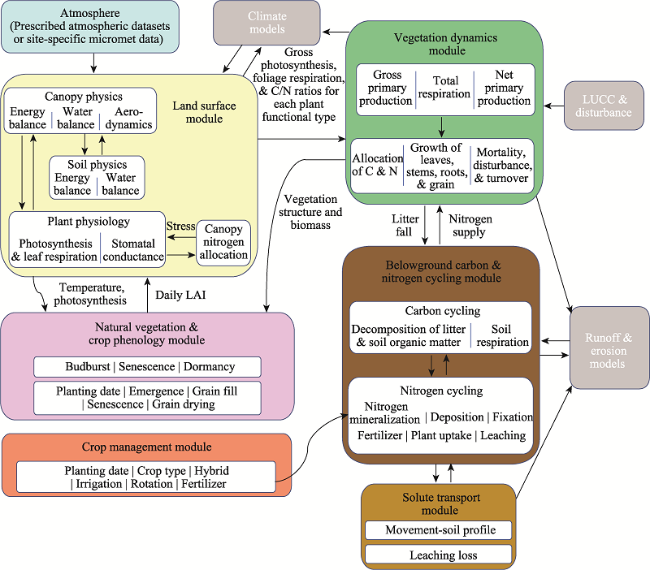
Fig. 1 IBIS key modules and their interactions (adopted and modified from Agro-IBIS) |
Table 1 Summary of IBIS model publications by study region, model name, ecosystem type, and research focus. |
| Application region | Number of publications | Model name | Number of publications | Ecosystem | Number of publications | Research focus* | Number of publications | |
|---|---|---|---|---|---|---|---|---|
| Canada-United States | 98 | IBIS | 177 | Multiple | 173 | Model Dev. | 52 | |
| Global | 70 | Agro-IBIS | 52 | Forest land | 93 | Hydrology | 36 | |
| China | 67 | CCM3-IBIS | 23 | Cropland | 68 | Vegetation | 25 | |
| Amazon/Brazil | 59 | GENESIS-IBIS | 9 | Grassland | 8 | NPP | 25 | |
| Africa | 22 | RegCM3-IBIS | 9 | Wetland | 4 | LUCC/Disturb. | 23 | |
| India | 19 | MRCB-IBIS | 9 | Savanna | 2 | Phenology | 11 | |
| Australia | 3 | TRIPLEX-GHG | 8 | Desert | 1 | Eco. Service | 9 | |
| Europe | 3 | Can-IBIS | 5 | Oil sand | 1 | Nitrogen loss | 6 | |
| Other | 16 | Other | - | Other | - | CH4/N2O | 5 | |
Note: *Summary of research focus is incomplete and represents only relative weights. NPP: Ecosystem net primary productivity; LUCC: Land use and land cover change. |
Work of Jinxun Liu and Carol Deering was funded by the U.S. Geological Survey Biologic Carbon Sequestration Assessment Program (LandCarbon).
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}