
Journal of Resources and Ecology >
The Facilitation of Restoration by Cushion Plant Androsace tapete in a Degraded Alpine Grassland
Received date: 2021-07-25
Accepted date: 2021-10-21
Online published: 2022-01-08
Supported by
The National Natural Science Foundation of China(31770477)
The Strategic Priority Research Program of the Chinese Academy of Sciences(XDA19050502)
The Strategic Priority Research Program of the Chinese Academy of Sciences(XDA20010201)
The National Key R&D Program of China(2017YFA0604801)
The National Key R&D Program of China(2016YFC0502001)
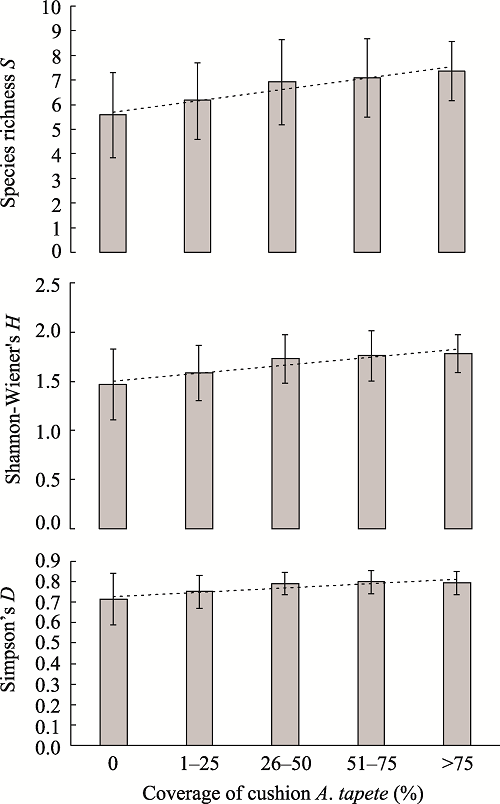
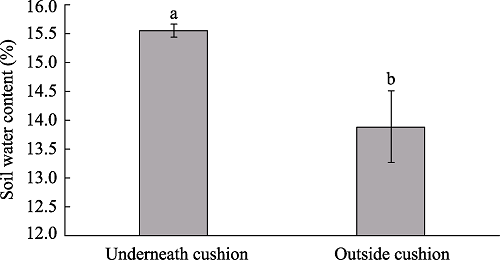
The cushion plant Androsace tapete is an endemic species that is widely distributed in the Qinghai-Tibetan Plateau, and also predominant in the alpine grassland that is locally degraded due to overgrazing and other reasons. As an ecosystem engineer cushion plant, its ability to facilitate the restoration of degraded alpine grassland was studied in a degraded alpine grassland at an elevation of 4500 m on the southern slope of the Nyainqentanglha Mountains in Damxung. The species diversity, soil nutrients and water content underneath and outside the cushion plant A. tapete were investigated. The results showed that soil nutrients underneath the A. tapete cushion were significantly increased by about 16%-48% compared to outside the cushion, of which the organic matter and total N were increased by 16.2% and 18.9% respectively, and the soil water content was increased about 12%. The index of species diversity of richness (S), Shannon-Wiener’s H and Simpson’s D all increased with the coverage of cushion plant A. tapete. Our results suggested that this cushion plant can facilitate restoration of the degraded alpine grassland by modifying the local soil environment and increasing the community diversity, so it should be conserved for the restoration of degraded alpine grasslands on the Qinghai-Tibetan Plateau.
HE Yongtao , WANG Fang , NIU Ben , WANG Zhipeng , LI Meng , SHI Peili , ZHANG Xianzhou . The Facilitation of Restoration by Cushion Plant Androsace tapete in a Degraded Alpine Grassland[J]. Journal of Resources and Ecology, 2022 , 13(1) : 107 -112 . DOI: 10.5814/j.issn.1674-764x.2022.01.012
Table 1 Community species composition in the sampled plots |
| Species | Coverage (%) | Species | Coverage (%) |
|---|---|---|---|
| Androsace tapete | 6.88 | Leontopodium sp. | 0.59 |
| Gentiana squarrosa | 6.40 | Kobresia pygmae | 0.53 |
| Artemisia desertorum | 4.58 | Stellera chamaejasme | 0.39 |
| Carex atrofusca | 4.06 | Koeleria cristata | 0.37 |
| Pleurospermum hookeri | 3.68 | Aster flaccidus | 0.33 |
| Potentilla nivea | 2.62 | Taraxacum sp. | 0.23 |
| Anaphalis sp. | 2.01 | Pedicularis kansuensis | 0.20 |
| Kobresia humilis | 1.37 | Veronica biloba | 0.17 |
| Poa sp. | 1.20 | Unidentified species | 0.16 |
| Stipa aliena | 1.20 | Kobresia robusta | 0.16 |
| Polygonum macrophyllum | 1.13 | Iris sp. | 0.09 |
| Silene repens | 1.01 | Allium sp. | 0.08 |
| Oxytropis sp. | 0.90 | Anemone obtusiloba | 0.08 |
| Androsace mariae | 0.89 | Thalictrum sp. | 0.08 |
| Stipa purpurea | 0.76 | Rhodiola sp. | 0.06 |
| Saussurea sp. | 0.60 | Microula sikkimensis | 0.04 |
Fig. 1 Species diversity increase with coverage of the cushion plant A. tapete |
Table 2 Soil nutrients underneath and outside the cushion plant A. tapete |
| Sample area | pH | Organic matter (g kg-1) | Total N (g kg-1) | NO3- (mg kg-1) | NH4+ (mg kg-1) | Available P (mg kg-1) | Available K (mg kg-1) | Slow-acting K (mg kg-1) |
|---|---|---|---|---|---|---|---|---|
| Underneath A. tapete | 6.26±0.06a | 28.93±6.78a | 1.49±0.23a | 0.24±0.03a | 1.40±0.21a | 5.72±1.72a | 68.52±12.99a | 463.31±78.70a |
| Outside A. tapete | 6.19±0.20a | 24.89±7.96b | 1.25±0.38b | 0.34±0.08b | 0.98±0.24b | 4.46±2.29a | 49.54±7.12b | 311.83±20.47b |
Note: Different letters (a, b) show the significant differences between underneath and outside the cushion plant A. tapete (P < 0.05). |
Fig. 2 Soil water contents underneath and outside the cushion plant A. tapete |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}