
Journal of Resources and Ecology >
Suppressive Effects of Traditional Mulching Using Japanese Knotweed (Fallopia japonica) on Solanaceae Crop Diseases
Received date: 2020-08-17
Accepted date: 2021-01-30
Online published: 2021-11-26
Supported by
Japan Society for the Promotion of Science (JSPS) KAKENHI(JP15K06930)
Japan Society for the Promotion of Science (JSPS) KAKENHI(JP18H02286)
Japan Society for the Promotion of Science (JSPS) KAKENHI(JP19K06108)
Poaceae plant species, such as silver grass, are commonly used in mulching activities Japan. In contrast, local farmers have traditionally used Japanese knotweed (Fallopia japonica) mulch in the cultivation of solanaceous crops in the Nishi-Awa area of Japan, which is a Globally Important Agricultural Heritage Systems site. We have previously evaluated the positive effects of Japanese knotweed mulching on solanaceous crops, such as eggplants, tomato, and potato. In the present study, we observed that the naturally occurring diseases in the solanaceous crops tended to decrease when the knotweed mulching system was adopted, in comparison to when Poaceae mulch was adopted. In eggplants, leaf mold and powdery mildew decreased under Japanese knotweed mulching. We further evaluated the effects of Japanese knotweed mulching by inoculating test plants with Pseudomonas cichorii. We observed suppression of bacterial disease and tomato mosaic virus under Japanese knotweed mulching and following spraying with Japanese knotweed extracts. In addition, disease-resistance genes were expressed at high levels in Arabidopsis thaliana, a model plant, following treatment with Japanese knotweed extracts. The results suggest that Japanese knotweed has potential applications in future sustainable agriculture activities.
INAGAKI Hidehiro , KUBOTA Sakiko , HASEGAWA Kana , UNNO Nahoko , USUI Yukiko , TAKIKAWA Yuichi . Suppressive Effects of Traditional Mulching Using Japanese Knotweed (Fallopia japonica) on Solanaceae Crop Diseases[J]. Journal of Resources and Ecology, 2021 , 12(6) : 869 -875 . DOI: 10.5814/j.issn.1674-764x.2021.06.014
Table 1 Effects of plant-mulching on plant disease in eggplant |
| Mulching treatments | 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Leaf mold (Mycovellosiella nattrassii) | Powdery mildew | Leaf mold (Mycovellosiella nattrassii) | |||||||
| No. of diseased plants | Lesion area percentage (%) | Preventive value | No. of diseased plants | Lesion area percentage (%) | Preventive value | No. of diseased plants | Lesion area percentage (%) | Preventive value | |
| Japanese knotweed mulching | 4/5 | 53.5 | 5.3 | 1/5 | 3.0 a | 75.0 | 4/5 | 11.0 a | 66.2 |
| Silver grass mulching | 4/5 | 49.5 | 10.5 | 1/5 | 3.0 a | 75.0 | 4/5 | 25.0 ab | 23.1 |
| Non-treatment | 4/5 | 59.5 | - | 4/5 | 12.0 b | - | 5/5 | 32.5 b | - |
| ANOVA | ns | ** | * | ||||||
Note: **, * indicate significant difference at 1% level and 5% level respectively in ANOVA after aricsin transform and ns indicates no siginificant difference. Different letters indicate significant differences among treatments based on Tukey’s multiple range test at 5% level. |
Table 2 Effects of plant-mulching on brown spot bacterial disease (Pseudomonas cichorii (Swingle) Stapp) in eggplant by inoculation test |
| Mulching treatments | Bacterial concentration (×10) | Bacterial concentration (×1) | ||
|---|---|---|---|---|
| Lesion area percentage (%) | Preventive value | Lesion area percentage (%) | Preventive value | |
| Japanese knotweed mulching | 24.2 a | 53.0 | 13.9 a | 62.9 |
| Silver grass mulching | 45.8 b | 11.1 | 28.7 ab | 23.5 |
| Non-treatment | 51.5 b | - | 37.5 b | - |
| ANOVA | ** | * | ||
Note: **, * indicate significant difference at 1% level and 5% level respectively in ANOVA after aricsin transform. Different letters indicate significant differences among treatments based on Tukey’s multiple range test at 5% level. |
Table 3 Effects of plant-mulching on plant disease in tomato |
| Mulching treatments | Tomato spotted bacterial disease (Pseudomonas syringae) | Tomato mosaic virus (ToMV) | |||||
|---|---|---|---|---|---|---|---|
| No. of diseased plants | No. of diseased leaves | Lesion area percentage (%) | Preventive value | No. of diseased plants | No. of diseased leaves | Preventive value | |
| Japanese knotweed mulching | 5/5 | 4.8 ab | 13.5 | 15.7 | 2/5 | 0.8 ab | 43.9 |
| Japanese knotweed extract | 5/5 | 3.6 a | 11.5 | 28.2 | 0/5 | 0 a | 100.0 |
| Non-treatment | 5/5 | 5.0 b | 16.0 | 0 | 3/5 | 1.4 b | 0 |
| ANOVA | * | ns | * | ||||
Note: * indicate significant difference at 5% level in ANOVA after aricsin transform, ns indicates no significant difference. Different letters indicate significant differences among treatments based on Tukey’ multiple range test at 5% level. |
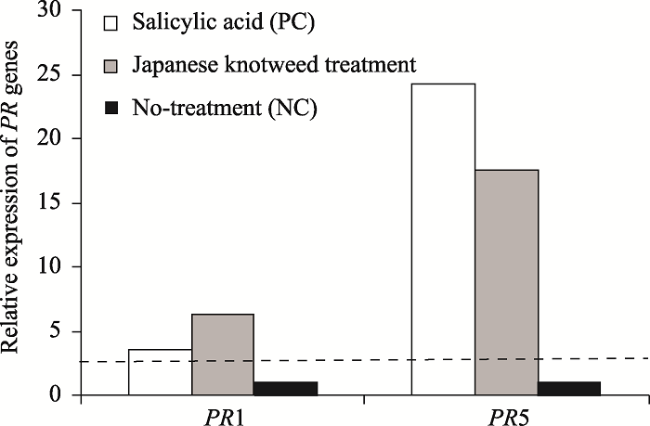
Fig. 1 Relative levels of expression of PR genes in Arabidopsis thaliana leaf under treatment with salicylic acid and Japanese knotweed extract |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
FAO Food Agriculture Organization. 2020a. Globally important agricultural heritage systems (GIAHS). 2020-08-06].
|
| [9] |
FAO Food Agriculture Organization. 2020b. Globally important agricultural heritage systems (GIAHS). Nishi-Awa Steep Slope Land Agriculture System.
|
| [10] |
|
| [11] |
|
| [12] |
GIAHS Book Editorial and
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
Syngenta. 2020. SAKALIA®. 2020-08-06].
|
| [42] |
|
| [43] |
|
| [44] |
TTGAHPC Tokushima Tsurugisan Globally Agricultural Heritage Promotion Council. 2019. Globally recognized Agriculture and Living in Nishi-Awa. 2020-08-06].
|
| [45] |
UNU United Nations University. 2011. Japanese agricultural heritage systems recognized. 2020-08-06].
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}