
Journal of Resources and Ecology >
Composition and Regeneration of Trees in the Community Forests of Lamjung District, Nepal
Received date: 2020-10-01
Accepted date: 2021-03-02
Online published: 2021-11-22
The community forest program in Nepal is one of the successful conservation initiatives. Tree species in a forest ecosystem have a fundamental role in maintaining the vegetation structure, complexity, and heterogeneity. This study analyzes the composition and regeneration of tree species in five community forests (CFs) of the sub-tropical region, Lamjung district, Nepal for preparing baseline data for long-term research projects and helps the community to manage their CFs scientifically. Tree species data were generated by stratified random sampling using 35 quadrate plots (size: 20 m x 20 m). The density of adult trees in the forests ranged from 575 Ind ha -1 (Tilahar CF) to 1196 Ind/ha (Deurali Thadopakha CF). The sapling ranged from 2533 Ind ha -1 (Tilahar CF) to 4000 Ind ha -1 (Thuliban CF) and seedling from 19583 Ind ha -1 (Tilahar CF) to 37500 Ind ha -1 (Thuliban CF). Similarly, the adult tree basal area varied from 28.34 m 2 ha -1 (Tilahar CF) to 49 m 2ha -1 (Deurali CF). The adult tree diversity index (Shannon-Weiner’s H) also ranged from 1.08 (Thuliban CF) to 1.88 (Tilahar CF). The tree species such as Sapium insigne, Ficus benghalensis, Lagerstroemia parviflore, Albizia sp. and Pinus roxburghii were weak regeneration. In general, the forests have good regeneration status except for Tilahar Community Forest, but based on the DBH size class distribution diagram, there is no sustainable regeneration. Among the five community forests the DBH size of adults was significantly different, and the DBH of Deurali Thadopakha was the lowest. But only few species have good regeneration and most of the species have weak, poor to no regeneration. The dominancy of fewer species like Shorea robusta, Castanopsis indica, and Schima wallichii accordingly maintain the overall regeneration of tree of CFs, so further plantation needs to be done inside the CF by triage accordingly those species whose regeneration has poor.
Key words: community forest; composition; regeneration; species; tree
Tilak Babu CHAPAGAI , Dipak KHADKA , Dinesh Raj BHUJU , Narendra Raj KHANAL , SHI Shi , CUI Dafang . Composition and Regeneration of Trees in the Community Forests of Lamjung District, Nepal[J]. Journal of Resources and Ecology, 2021 , 12(5) : 658 -668 . DOI: 10.5814/j.issn.1674-764x.2021.05.009
Table 1 General characteristics of the community forests (CF) selected for study in Lamjung District, Nepal. |
| Name of CF | Area (ha) | Altitude (m) | Latitude | Longitude | Dominant species |
|---|---|---|---|---|---|
| Satipatal (SCF) | 66.77 | 807 | 28°10°39.15N | 84°26°41.87E | Shorea robusta |
| Thuliban (TBCF) | 24.22 | 803 | 28°11°15.90 N | 84°26°8.51E | Shorea robusta |
| Deurali (DCF) | 57.67 | 887 | 28°11°47.08 N | 84°25°46.66E | Castanopsis indica |
| Deurali Thadopakha (DTCF) | 19.34 | 1114 | 28°12°36.82 N | 84°26°18.74E | Castanopsis indica |
| Tilahar (TCF) | 44.11 | 1140 | 28°12°27.49N | 84°26°13.39E | Schima wallichii |
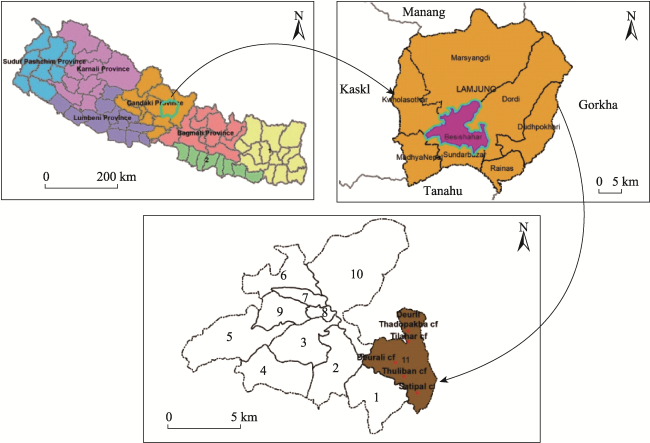
Fig. 1 Map of the study area showing studied community forest of Lamjung District,NepalNote: The numbers from 1-11 are the respective wards of Besishahar Municipality. |
Table 2 The number of recorded species |
| Serial number | Name of CFs | Number of adult | Number of sapling | Number of seedling |
|---|---|---|---|---|
| 1 | Satipatal | 432 | 144 | 113 |
| 2 | Thuliban | 195 | 100 | 75 |
| 3 | Deurali | 344 | 134 | 88 |
| 4 | Deurali Thadopakha | 287 | 110 | 67 |
| 5 | Tilahar | 138 | 76 | 47 |
Table 3 Community Structure of trees in community forests |
| Indices | Tree layer | SCF | TBCF | DCF | DTCF | TCF | Average |
|---|---|---|---|---|---|---|---|
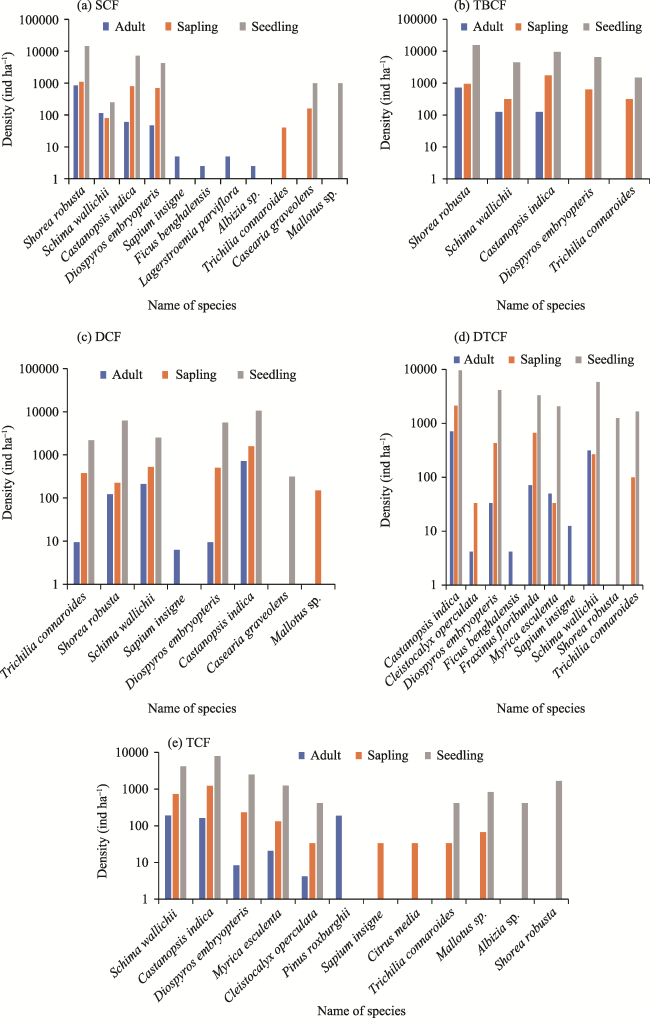
| Density (Ind ha-1) | Sapling | 2880 | 4000 | 3350 | 3667 | 2533 | 3286 |
| Seedling | 28250 | 37500 | 27500 | 27917 | 19583 | 28150 | |
| Basal Area/Abundance (BA/A) | Adult | 42.50 | 46.80 | 49.00 | 37.73 | 28.34 | 40.87 |
| Sapling | 17.60 | 14.40 | 15.69 | 19.17 | 19.40 | 17.25 | |
| Seedling | 10.41 | 10.73 | 8.62 | 19.37 | 14.06 | 12.64 | |
| Shannon-Weiner’s | Adult | 1.17 | 1.08 | 1.37 | 1.66 | 1.88 | 1.43 |
| diversity index (H) | Sapling | 2.00 | 2.02 | 2.16 | 1.81 | 2.03 | 2.00 |
| Seedling | 1.81 | 2.02 | 2.16 | 2.50 | 2.49 | 2.20 | |
| Simpson’s diversity | Adult | 0.38 | 0.41 | 0.50 | 0.57 | 0.70 | 0.51 |
| index (D) | Sapling | 0.71 | 0.71 | 0.71 | 0.61 | 0.67 | 0.68 |
| Seedling | 0.65 | 0.71 | 0.74 | 0.79 | 0.76 | 0.73 | |
| Species richness (S) | 11 | 5 | 8 | 10 | 12 | 9 |
Notes: SCF= Satipatal Community Forest; TBCF= Thuliban Community Forest; DCF=Deurali Community Forest; DTCF= Deurali Thadopakha Community Forest; TCF= Tilahar Community Forest. |
Fig. 2 Adult, sapling, and seedling densities in different communities forests (CFs) of Lamjung District, Nepal. |
Table 4 Species-IVI value in different CFs |
| Name of species | SCF | TBCF | DCF | DTCF | TCF | Average |
|---|---|---|---|---|---|---|
| Albizia sp. | 3.58 | 0 | 0 | 0 | 0 | 0.72 |
| Castanopsis indica | 30.52 | 50.32 | 122.43 | 119.29 | 67.26 | 77.96 |
| Cleistocalyx operculate | 0 | 0 | 0 | 5.70 | 7.15 | 2.57 |
| Diospyros embryopteris | 22.67 | 0 | 9.07 | 13.29 | 9.50 | 10.91 |
| Ficus benghalensis | 3.58 | 0 | 0 | 6.49 | 0 | 2.01 |
| Fraxinus floribunda | 0 | 0 | 0 | 17.78 | 0 | 3.56 |
| Lagerstroemia parviflora | 3.89 | 0 | 0 | 0 | 0 | 0.78 |
| Myrica esculenta | 0 | 0 | 0 | 20.80 | 24.91 | 9.14 |
| Pinus roxburghii | 0 | 0 | 0 | 0 | 79.41 | 15.88 |
| Sapium insigne | 4.03 | 0 | 8.31 | 11.10 | 0 | 4.69 |
| Schima wallichii | 44.34 | 64.62 | 92.53 | 115.08 | 111.76 | 85.67 |
| Shorea robusta | 187.39 | 185.06 | 58.99 | 0 | 0 | 86.29 |
| Trichilia connaroides | 0 | 0 | 8.67 | 0 | 0 | 1.73 |
Notes: SCF= Satipatal Community Forest; TBCF= Thuliban Community Forest; DCF=Deurali Community Forest; DTCF= Deurali Thadopakha Community Forest; TCF= Tilahar Community Forest. |
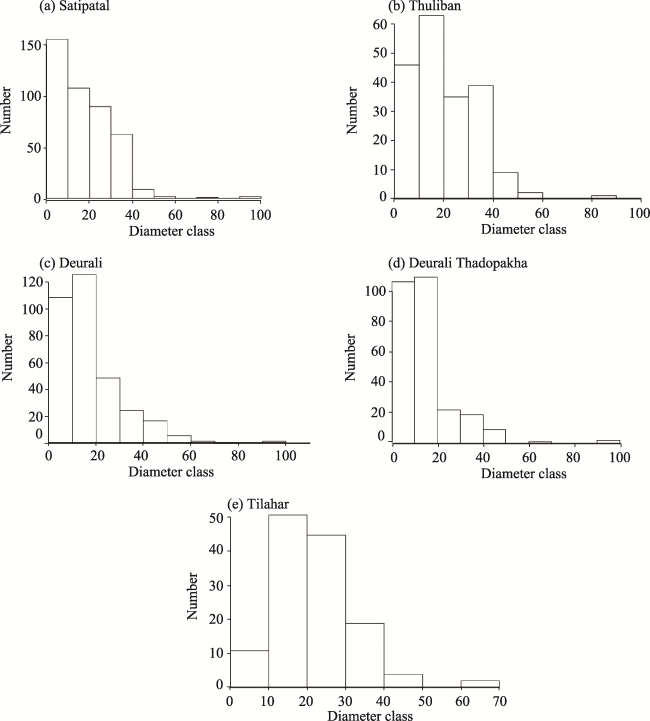
Fig. 3 Population structure of adults of different community forests of Lamjung, number of trees recorded in Y axis (Number) and diameter at breast height (DBH) in centimeter (cm) (Diameter class) in X axis. |
Table 5 Summary of DBH in different forests of Lamjung |
| Parameterss | SCF | TBCF | DCF | DTCF | TCF |
|---|---|---|---|---|---|
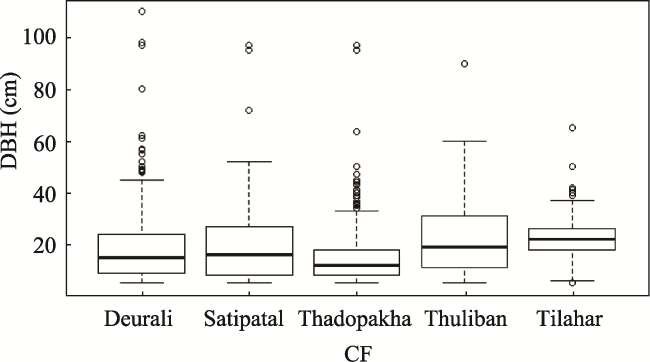
| Mean | 27.00 | 31.00 | 19.09 | 15.80 | 26.00 |
| Max. | 97.00 | 90.00 | 110.00 | 97.00 | 65.00 |
| Min | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Q1 | 8.00 | 11.00 | 9.00 | 8.07 | 18.00 |
| Q2 | 16.00 | 19.00 | 15.00 | 12.00 | 22.00 |
| Q3 | 18.90 | 31.00 | 24.00 | 18.00 | 26.00 |
| Bowely coefficient of skewness | -0.47 | 0.20 | 0.20 | 0.21 | 0.00 |
Note: SCF= Satipatal Community Forest; TBCF= Thuliban Community Forest; DCF=Deurali Community Forest; DTCF= Deurali Thadopakha Community Forest and TCF= Tilahar Community Forest. |
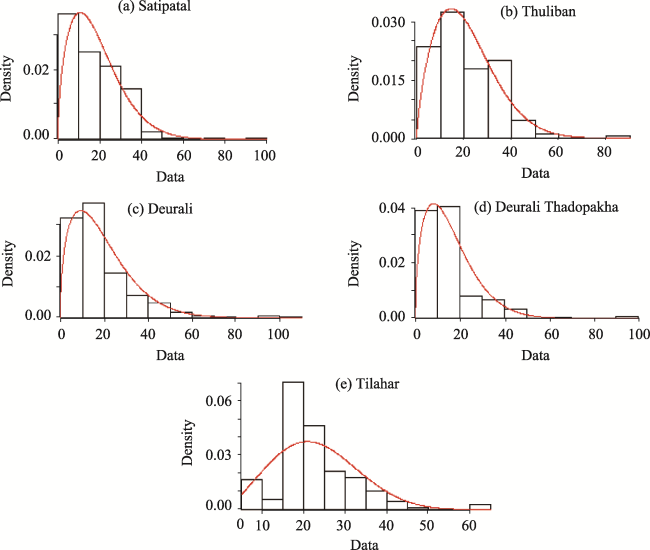
Table 6 Shape and scale parameters (with standard errors) of Weibull distribution for different CFs indicated Weibul distribution of DBH of trees of all the five studied forests are in mound shaped |
| Community Forest | Weibul shape parameter (Standard errors ) | Weibul scale parameter (Standard error) |
|---|---|---|
| Satipatal | 1.56 (0.06) | 20.69 (0.68) |
| Thuliban | 1.74 (0.09) | 23.78 (1.04) |
| Deurali | 1.43 (0.06) | 21.21 (0.86) |
| Deurali Thadopakha | 1.57 (0.07) | 18.93 (0.73) |
| Tilahar | 2.42 (0.15) | 25.95 (0.98) |
Note: The number in the bracket are standard error. |
Fig. 5 Comparison of diameter at breast height (DBH) in centimeter (cm) size class distribution of trees in different community forests (CF) of Lamjung District, Nepal. |
Fig. 4 Probability Density Function Weibul in Y axis for Weibull Distribution plotted with data (DBH classes of trees in cm) of studied forests in X axis starting from Satipatal followed by Thuliban, Deurali, Deurali Thadopakha, and Tilahar respectively from the top left side. |
Table 7 Species regeneration status of community forests |
| Name of species | Local name | Family | SCF | TBCF | DCF | DTCF | TCF |
|---|---|---|---|---|---|---|---|
| Shorea robusta | Sal | Dipterocarpaceae | G | G | G | N | N |
| Schima wallichii | Chilaune | Theaceae | F | G | G | F | G |
| Castanopsis indica | Katush | Fagaceae | G | G | G | G | G |
| Diospyros embryopteris | Tindu | Ebeniaceae | G | N | G | G | G |
| Sapium insigne | Khirro | Euphorbiaceae | NO | ― | NO | NO | N |
| Ficus benghalensis | Bar | Moraceae | NO | ― | ― | NO | ― |
| Lagerstroemia parviflora | Bot dhayaro | Lythraceae | NO | ― | ― | ― | ― |
| Albizia sp. | Siris | Leguminosae | NO | ― | ― | ― | N |
| Trichilia connaroides | Ankhatare | Meliaceae | N | N | G | N | N |
| Casearia graveolens | Badkamlo | Salicaceae | N | ― | N | ― | ― |
| Mallotus sp. | Sindhure | Euphorbiaceae | N | ― | N | ― | N |
| Cleistocalyx operculate | Kyamuna | Myrtaceae | ― | ― | ― | P | G |
| Fraxinus floribunda | Lankure | Oleaceae | ― | ― | ― | G | ― |
| Myrica esculenta | Kaphal | Myricaceae | ― | ― | ― | F | G |
| Citrus media | Bimiro | Rutaceae | ― | ― | ― | ― | NO |
| Pinus roxburghii | Salla | Pinaceae | ― | ― | ― | ― | N |
Note: G = Good, F = Fair, P = Poor, N = New, NO = None, ― = Absence of given species; SCF= Satipatal Community Forest; TBCF= Thuliban Community Forest; DCF=Deurali Community Forest; DTCF= Deurali Thadopakha Community Forest; TCF= Tilahar Community. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
DFO. 2016. Community forest user group monitoring and assessment. Lamjung, Nepal: District Forest Office.
|
| [12] |
DoF. 2014. Community Forest Database. Kathmandu, Nepal: Department of Forests (DoF).
|
| [13] |
FRA. 2015. Middle mountains forests of Nepal. Department of forest research and survey ministry of forests and soil conservation government of Nepal.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}