
Journal of Resources and Ecology >
Study of Vulture Habitat Suitability and Impact of Climate Change in Central India Using MaxEnt
Received date: 2020-06-16
Accepted date: 2020-08-15
Online published: 2021-03-30
Vultures provide invaluable ecosystem services and play an important role in ecosystem balancing. The number of native vultures in India has declined in the past. Acquiring present knowledge of their habitat spread is essential to manage and prevent such a decline. It is envisaged that ongoing climate crisis may further cause change in habitat suitability and impact the existing population. Therefore, this study in Central India—a vulture stronghold, is aimed at predicting habitat changes in the short and long term and present the data statistically and graphically by using Species Distribution Model. MaxEnt software was chosen for its advantages over other models, like using presence-only data and performing well with incomplete data, small sample sizes and gaps, etc. Global Climate Model ensemble (CCSM4, HadGEM2AO and MIROC5), was used to get better prediction. Fourteen robust models (AUC 0.864-0.892) were developed using data from over 1000 locations of seven vulture species over two seasons together. Selected climatic and other environmental variables were used to predict the current habitat. Future prediction was based on climatic variables only. The most important variables influencing the distribution were precipitation (bio 15, bio 18, bio 19) and temperature (bio 3, bio 5). Forest and water bodies were the major influencers within land use-landcover in the current prediction. At finer scale, while extremely suitable habitat area decreased and highly suitable area increased over time, the total suitable area marginally increased in 2050 but decreased in 2070. For broader consideration, net loss in suitable area was 5% in 2050 and 7.17% in 2070 (RCP4.5). Similarly, in the RCP8.5 this was 6% in 2050 and 7.3% in 2070. The data generated can be used in conservation planning and management and thus protecting the vultures from any future threat.
Kaushalendra K. JHA , Radhika JHA . Study of Vulture Habitat Suitability and Impact of Climate Change in Central India Using MaxEnt[J]. Journal of Resources and Ecology, 2021 , 12(1) : 30 -42 . DOI: 10.5814/j.issn.1674-764x.2021.01.004
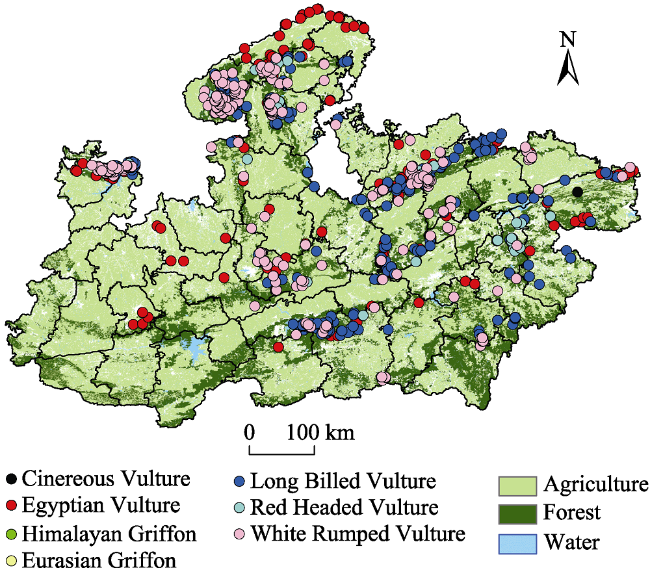
Fig. 1 Location of vulture species in Central India surveyed in 2016Note: This presence only record has been used in Species Distribution Model, MaxEnt, as sample input. Proximity of vulture locations may be noted to forested landscape in most cases. |
Table 1 Selected variables used in vulture habitat prediction model |
| Bio-climatic | Environmental | |
|---|---|---|
| Temperature | Precipitation | |
| (i) bio 3 = Isothermality (bio 2/bio 7) ×100 | (v) bio 13 = Precipitation of wettest month | (x) LULC (Forest, water, rural and urban built-up area, agriculture, wasteland, scrubland) |
| (ii) bio 5 = Max temperature of warmest month | (vi) bio 14 = Precipitation of driest month | (xi ) NDVI (January 2016) (xii) NDVI (May 2016) |
| (iii) bio 9 = Mean temperature of driest quarter | (vii) bio 15 = Precipitation seasonality (coefficient of variation) | (xiii) Elevation |
| (iv) bio 11 = Mean temperature of coldest quarter | (viii) bio 18 = Precipitation of warmest quarter | |
| (ix) bio 19 = Precipitation of coldest quarter | ||
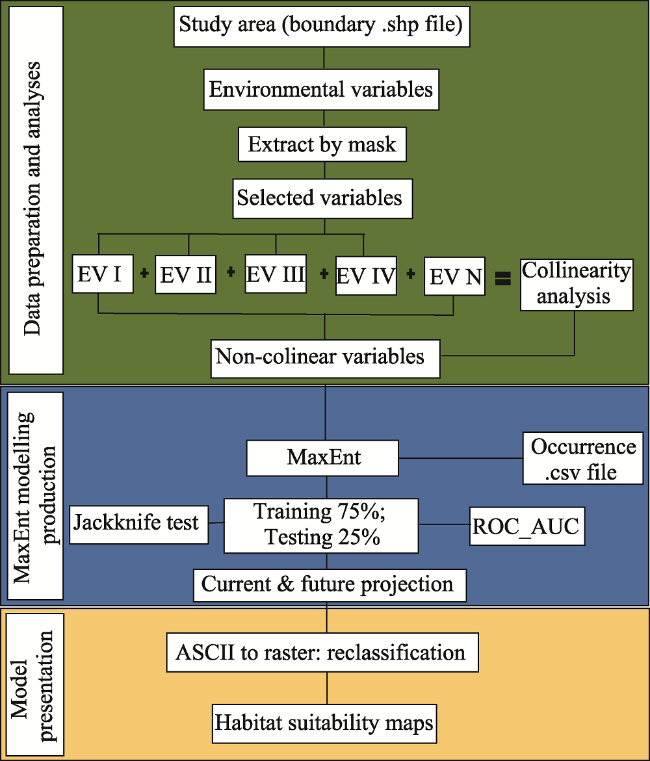
Fig. 2 Flow chart of Species Distribution Modelling using MaxEnt softwareNote: EV I to EV IV are environmental variables (Table 1); N indicates any number of EV. Flow is top to bottom along connectors. |
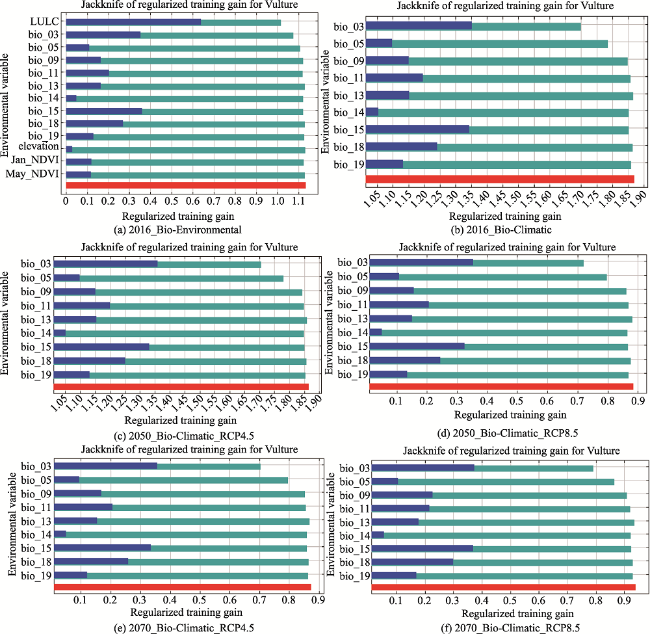
Fig. 3 Jackknife charts of variable contribution in model predictionNote: The bars in light blue represent without variable; the bars in dark blue represent with only variable; the bars in red represent with all variables. |
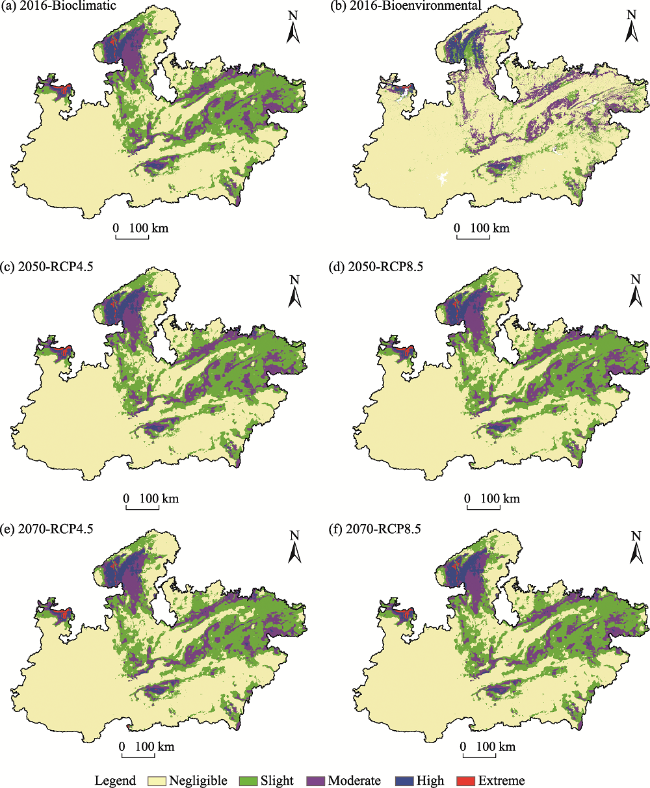
Fig. 4 The expanse of habitat suitability classes under different scenariosNote: Top two maps compare impact of LULC inclusion in terms of reduced suitable area in the top right map. |
Table 2 Habitat suitability area (km2; average of three GCMs) distribution among different categories under varied scenario |
| Habitat suitability category | Current (2016) | Short term (2050) | Long term (2070) | |||
|---|---|---|---|---|---|---|
| Environmental | Climatic | Climatic | Climatic | |||
| RCP4.5 | RCP8.5 | RCP4.5 | RCP8.5 | |||
| Least suitable | 242265 (79.3) | 189013 (61.1) | 187442 (60.6) | 188833 (61.0) | 191174 (61.8) | 190816 (61.7) |
| Slightly suitable | 27733 (9.1) | 78313 (25.3) | 79883 (25.8) | 79138 (25.6) | 76827 (24.8) | 78066 (25.2) |
| Moderately suitable | 28089 (9.2) | 33930 (11.0) | 33347 (10.8) | 33330 (10.8) | 32513 (10.5) | 31633 (10.2) |
| Highly suitable | 7310 (2.4) | 7336 (2.4) | 8020 (2.6) | 7357 (2.4) | 8139 (2.6) | 8118 (2.6) |
| Extremely suitable | 217 (0.1) | 829 (0.3) | 729 (0.2) | 764 (0.2) | 769 (0.2) | 788 (0.3) |
Note: Figures in parentheses are percentage of the total area available. |
Table 3 Data representing the expanse of habitat suitability and possible future change in km2 |
Category | Bioclimatic | RCP4.5 | RCP8.5 | ||
|---|---|---|---|---|---|
| 2016 | 2050 | 2070 | 2050 | 2070 | |
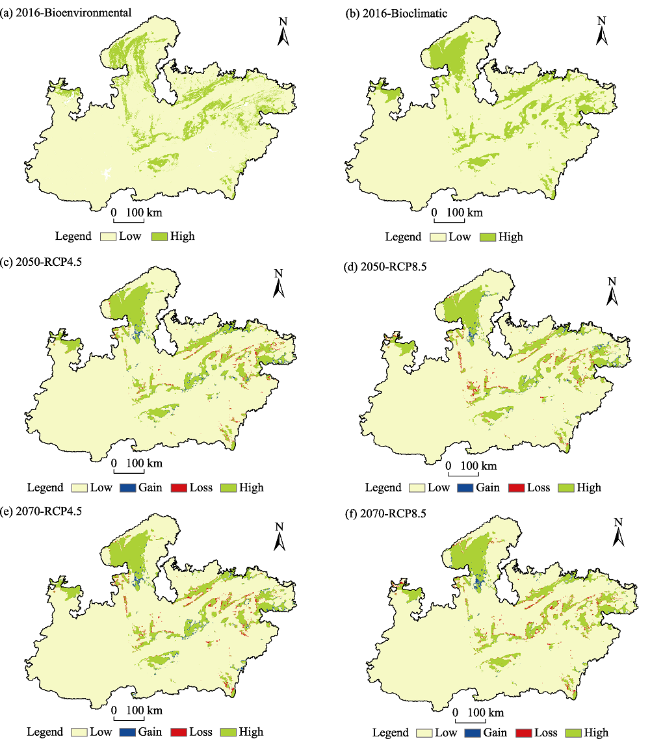
| Low habitat suitability | 266641 | 264391 | 264294 | 264599 | 265111 |
| Gain | 0 | 2250 | 2347 | 2042 | 1530 |
| Loss | 0 | 2249 | 3020 | 2685 | 3082 |
| High habitat suitability | 41987 | 39738 | 38967 | 39302 | 38905 |
Fig. 5 The changes in vulture habitat area suitability from low to high and vice versa in short term (2050) and long term (2070) with respect to present (2016) |
Authors are thankful to the Forest Department and Biodiversity Board, Madhya Pradesh in supporting vulture count in 2016. Dr. Advait Edgaonkar, Assistant Professor and Miss Amreesh Bhullar, Technical Assistant, Geoinformatics Laboratory, IIFM, Bhopal deserve appreciation for their contribution and support.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
Anon. 2004. Report of the international south Asian vulture recovery plan workshop. Parwanoo, India.
|
| [8] |
Anon. 2006. Action plan for vulture conservation in India. Ministry of Environment and Forests, Government of India.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}