
Journal of Resources and Ecology >
Species Diversity Characteristics of a Natural Pinus taiwanensis Community with Different Diameter Classes and Forest Densities
|
WANG Yang, E-mail: 13296698026@163.com |
Received date: 2019-12-11
Accepted date: 2020-03-02
Online published: 2020-09-30
Supported by
Scientific Research Project of Education Department of Hubei Province(B2018481)
Scientific Research Project of Education Department of Hubei Province(B2018482)
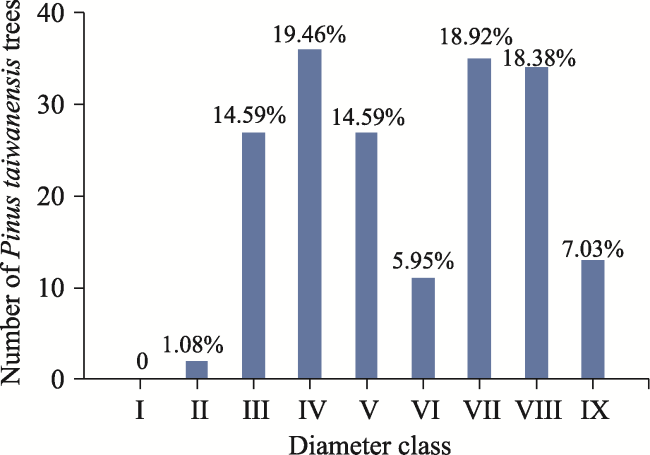
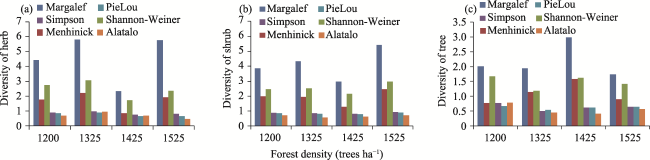
Pinus taiwanensis is a species endemic to China. This study selected four typical plots of Pinus taiwanensis in the natural secondary forest around Macheng City, in order to reveal the characteristics of and the relationships between different diameter classes (determined based on the diameter at breast height or DBH), forest densities and species diversity, as well as the similarities of species diversity of different plots within the community. The result showed that Pinus taiwanensis was the dominant species in the community. The ratio of Pinus taiwanensis trees of diameter class IV reached a peak of 19.46% of the total followed by diameter class VII at 18.92%. The study recorded 156 species of vascular plants from 130 genera of 71 families; Pinus taiwanensis was the dominant species in the community. When the forest density was 1200 trees ha -1 with the largest average diameter of DBH=36.779±4.444 cm, the diversity (Shannon index H'=1.6716) and the evenness (Pielou index E=0.6727) of the tree layer was the highest. When the forest density reached 1525 trees ha -1 with the lowest average diameter of DBH=18.957±5.141 cm, the richness (Dma=5.4308), the diversity (H'=2.9612) and the evenness (E=0.8985) of all shrub layers climbed to the maximum. When the forest density was 1325 trees ha -1, the richness (Dma=5.8132), the diversity (H'=3.0697) and the evenness (E=0.9025) of all herb layers peaked. In terms of vertical structure, the average diversity indexes were herb layer>shrub layer>tree layer. High canopy density weakened light intensity in the community, causing a reduction in the species diversities of herbs and shrubs. The average similarity coefficient between the sample plots was 0.3356, which was at the medium dissimilarity level. External disturbances and improper management were major contributors to the low species diversity of the community. The implementation of scientific management measures is urgently needed to optimize the forest structures of Pinus taiwanensis, create a benign community environment, and promote species diversities and establish a stable forest community structure.
Key words: Pinus taiwanensis; diameter class; forest density; species diversity
WANG Yang , JIANG Xiongbo , WU Dezhi . Species Diversity Characteristics of a Natural Pinus taiwanensis Community with Different Diameter Classes and Forest Densities[J]. Journal of Resources and Ecology, 2020 , 11(4) : 349 -357 . DOI: 10.5814/j.issn.1674-764x.2020.04.003
Table 1 General status of sample plots of natural Pinus taiwanensis forests in Macheng |
| Plot No. | Elevation (m) | Longitude | Latitude | Slope (˚) | Aspect | Canopy density | Forest density (trees ha-1) | Average DBH (cm) | Average height (m) | Average biomass (t ha-1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 792 | 115°17′23″ | 31°22′34″ | 20 | Southeast | 0.85 | 1425 | 22.479±10.114 | 13.913±3.942 | 296.097 |
| 2 | 812 | 115°18′50″ | 31°22′56″ | 15 | North | 0.70 | 1525 | 18.957±5.141 | 11.502±2.206 | 197.714 |
| 3 | 739 | 115°20'21 | 31°23'32 | 22 | Northeast | 0.65 | 1325 | 29.470±8.694 | 11.775±3.468 | 297.193 |
| 4 | 658 | 115°21′41″ | 31°23′25″ | 18 | Southeast | 0.80 | 1200 | 36.779±4.444 | 14.865±1.828 | 664.605 |
Note: 1, 2, 3 and 4 designate the four sample plots and this numbering will be used in the tables that follow. |
Fig. 1 The ratio of Pinus taiwanensis trees in different diameter class |
Table 2 Species diversity of natural Pinus taiwanensis communities at different forest densities |
| Plot No. | Vegetation layer | Richness | Diversity | Evenness | ||||
|---|---|---|---|---|---|---|---|---|
| Patrick (S) | Margalef (Dma) | Menhinick (Dmc) | Shannon-Weiner (H') | Simpson GINI (D) | PieLou (E) | Alatalo (Ea) | ||
| 1 | Herb | 14 | 2.3267 | 0.8568 | 1.7314 | 0.7618 | 0.6561 | 0.6879 |
| Shrub | 16 | 2.9666 | 1.2769 | 2.1661 | 0.8266 | 0.7812 | 0.6170 | |
| Tree | 14 | 2.9839 | 1.5852 | 1.6215 | 0.6203 | 0.6144 | 0.4023 | |
| 2 | Herb | 34 | 5.7461 | 1.9249 | 2.3669 | 0.8237 | 0.6712 | 0.4836 |
| Shrub | 27 | 5.4308 | 2.4648 | 2.9612 | 0.9306 | 0.8985 | 0.7314 | |
| Tree | 9 | 1.7410 | 0.9045 | 1.4145 | 0.6369 | 0.6438 | 0.5631 | |
| 3 | Herb | 32 | 5.8132 | 2.2242 | 3.0697 | 0.9853 | 0.9025 | 0.9593 |
| Shrub | 22 | 4.3351 | 1.9522 | 2.5248 | 0.8673 | 0.8168 | 0.5687 | |
| Tree | 9 | 1.9384 | 1.1430 | 1.1886 | 0.5016 | 0.5410 | 0.4408 | |
| 4 | Herb | 24 | 4.4244 | 1.7839 | 2.7411 | 0.9097 | 0.8625 | 0.6944 |
| Shrub | 18 | 3.8577 | 1.9878 | 2.4750 | 0.8876 | 0.8563 | 0.7253 | |
| Tree | 12 | 2.0040 | 0.7714 | 1.6716 | 0.7705 | 0.6727 | 0.7768 | |
Fig. 2 Species diversities at different forest densities |
Table 3 The importance value of tree, shrub and herb species in the 4 sample plots |
| Plot No. | Tree species | IVt(%) | Shrub species | IVs(%) | Herb species | IVh(%) |
|---|---|---|---|---|---|---|
| 1 | Pinus taiwanensis | 84.33 | Rhododendron simsii | 34.72 | Oplismenus undulatifolius | 39.33 |
| Paulownia fortunei | 2.17 | Camellia sinensis | 21.74 | Goodyera repens | 18.73 | |
| Vernicia fordii | 2.07 | Euscaphis japonica | 8.39 | Alpinia japonica | 15.73 | |
| Diospyros lotus | 1.72 | Wisteria sinensis | 6.63 | Aristolochia mollissima | 14.61 | |
| Euscaphis japonica | 1.57 | Alangium platanifolium | 5.19 | Viola verecunda | 3.75 | |
| Celtis biondii | 0.78 | Viburnum dilatatum | 4.40 | Parthenocissus dalzielii | 2.25 | |
| 2 | Pinus taiwanensis | 81.84 | Lindera erythrocarpa | 22.31 | Polygonum hydropiper | 34.94 |
| Carya cathayensis | 5.60 | Lindera glauca | 10.65 | Oplismenus undulatifolius | 19.23 | |
| Quercus serrata | 1.68 | Euscaphis japonica | 6.92 | Alpinia japonica | 8.33 | |
| Symplocos paniculata | 1.16 | Schisandra sphenanthera | 5.90 | Viola verecunda | 4.81 | |
| Mallotus japonicus | 0.70 | Corylus heterophylla | 5.88 | Teucrium viscidum | 4.49 | |
| Lindera erythrocarpa | 0.59 | Phyllanthus glaucus | 4.27 | Phryma leptostachya | 4.81 | |
| 3 | Pinus taiwanensis | 81.12 | Rhododendron simsii | 38.61 | Geum aleppicum | 13.43 |
| Lindera glauca | 6.69 | Lindera erythrocarpa | 12.87 | Kummerowia striata | 14.63 | |
| Dalbergia hupeana | 4.48 | Camellia sinensis | 11.88 | Erigeron annuus | 10.79 | |
| Carya cathayensis | 3.57 | Akebia trifoliata | 8.91 | Setaria faberii | 8.87 | |
| Diospyros lotus | 2.21 | Kerria japonica | 6.93 | Crassocephalum crepidioides | 6.71 | |
| Castanea seguinii | 1.76 | Lespedeza formosa | 5.94 | Fragaria nilgerrensis | 5.04 | |
| 4 | Pinus taiwanensis | 63.10 | Lespedeza formosa | 15.85 | Anaphalis sinica | 20.44 |
| Quercus acutissima | 15.79 | Lespedeza buergeri | 15.23 | Aster ageratoides | 12.71 | |
| Quercus serrata | 12.26 | Rhododendron simsii | 14.63 | Amphicarpaea edgeworthii | 8.84 | |
| Dalbergia hupeana | 4.20 | Glochidion wilsonii | 8.54 | Syneilesis aconitifolia | 8.29 | |
| Albizia kalkora | 2.31 | Elaeagnus pungens | 4.54 | Fragaria nilgerrensi | 5.65 | |
| Quercus variabilis | 1.32 | Rosa cymosa | 3.95 | Cyperus rotundus | 5.52 |
Table 4 The diversity similarity indexes of 4 plots in the Pinus taiwanensis community |
| Plot No. | 1 | 2 | 3 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jaccard (Cj) | Sǿrenson(Cs) | Ochiai(Co) | Jaccard (Cj) | Sǿrenson(Cs) | Ochiai(Co) | Jaccard (Cj) | Sǿrenson(Cs) | Ochiai(Co) | |||||||
| 2 | 0.3412 | 0.5088 | 0.3436 | ||||||||||||
| 3 | 0.2022 | 0.3364 | 0.2540 | 0.3232 | 0.4886 | 0.3284 | |||||||||
| 4 | 0.1951 | 0.3265 | 0.2469 | 0.2680 | 0.4228 | 0.2982 | 0.3295 | 0.4957 | 0.3319 | ||||||
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
Editorial Committee of Flora Reipublicae Popularis Sinicae, Chinese Academy of Sciences. 1978. Flora Reipublicae Popularis Sinicae. Beijing: Science Press, 266. (in Chinese)
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}