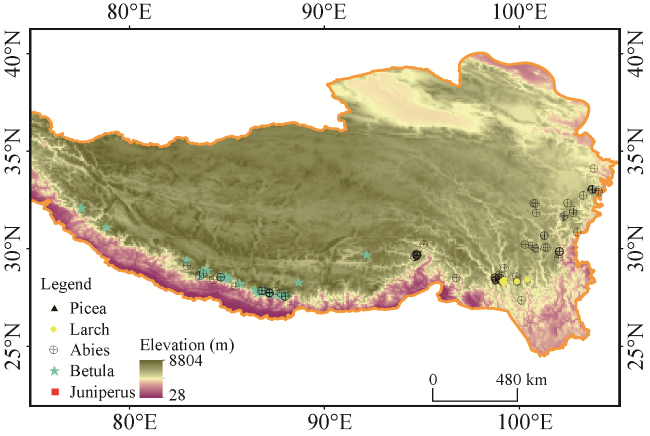
Spruce-fir forests are distributed mainly in the southeastern part of the Tibetan Plateau, extending between 85-105°E and 26-38°N. In the southeastern part of the Tibetan Plateau and southern slope of the Himalayan Range, the coniferous species are diverse. It is reported that there are 16 species of
Abies, 16 species of
Picea, six species of
Larix and 11 species of
Juniperus (
Sabina) on the Plateau. But only 14 species of
genus Abies, five of
Picea, five of
Juniperus and four of
Larix can reach climatic forest limit and consist of timberline species. In addition, schlerphyllous
Quercus and deciduous broad-leaved trees, e.g.
Betula, etc. can also form forest limit vegetation in west Himalaya, southeast Tibet and north Hengduan Mountaints. For example,
Betula utilis presents at treeline in Uttarakhand, India. The Hengduan Mts. serve as species differentiation center of
Abies. There are nine species of
Abies in this region, of which
Abies ferreana,
A. squamata,
A. nukiangensis,
A. delavayi,
A. georgei,
A. georgei var.
smithii, and
A. forrestii constitue forest limit species. Kangding, in western Sichuan Province, is the species differentiation center of genus
Picea and there covers over 10 species of spruce forests of the Qinghai-Tibet Plateau. But only some species such as
Picea balfouriana,
P. purpurea,
P. likiangensis and
P. crassifolia can reach forest limit and form timberline, in which
P. balfouriana is the most common and widely distributed timberline species in eastern Tibet and northwestern Sichuan plateau.
Larix is the forest limit genus on sunny slopes. In western Sichuan,
L. potaninii is the widespread timberline species and can extend to Bailongjiang watershed of southern Gansu Province.
L. potanini var.
marcrocarpa is widely distributed in southwestern Sichuan, northwestern Yunnan and northeastern Tibet. At the elevation of 3800-4300 m, forest limit forms together with
Aibes.
Sabina (now
Juniperus) is the alpine tree species and is the highest forest type to constitute forest limit.
S. convallium,
S. sultuaria and
S. tibetica are the timberline species on sunny slopes in western Sichuan and eastern Tibet. In eastern Qinghai, southern Gansu and northwestern Sichuan,
S. przewalskii, and
S. komarovii are dominant timberline species on sunny slopes (
Fig. 1).

{kind=link}
{kind=link}
{kind=link}
{kind=link}