
Journal of Resources and Ecology >
A Meta-analysis of the Effects of Organic and Inorganic Fertilizers on the Soil Microbial Community
|
WANG Jiangwei, E-mail: wangjw.15s@igsnrr.ac.cn |
Received date: 2019-12-12
Accepted date: 2020-02-08
Online published: 2020-06-16
Supported by
The National Key Research and Development Program of China(2016YFC0502001)
The National Key Research and Development Program of China(2016YFC0502005)
The National Key Research and Development Program of China(2017YFA0604801)
The Tibet Natural Science Foundation(XZ 2018 ZR G-141)
(31600432)
The National Natural Science Foundation of China(31600432)
(41571042)
The National Natural Science Foundation of China(41571042)
The Tibet Science and Technology Major Projects of the Pratacultural Industry.
Copyright
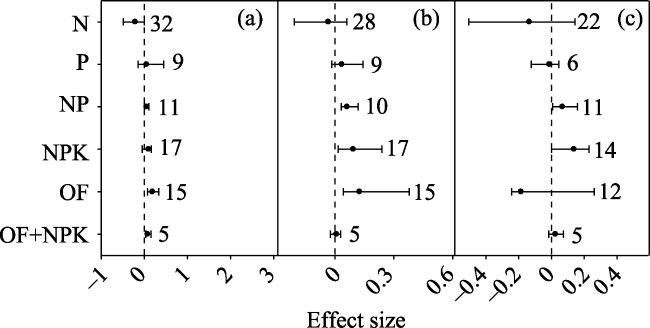
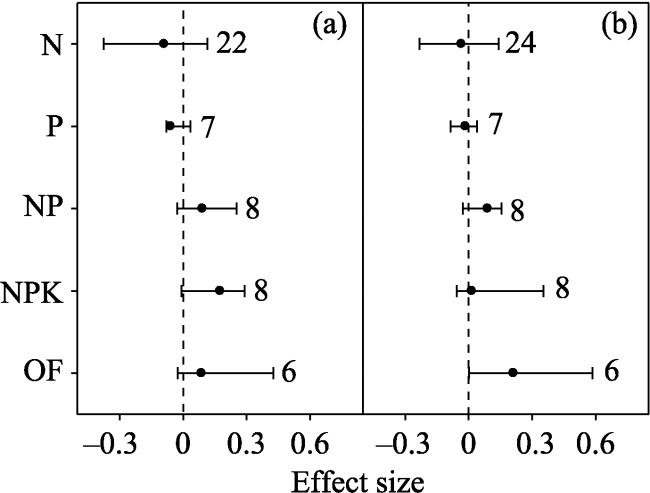
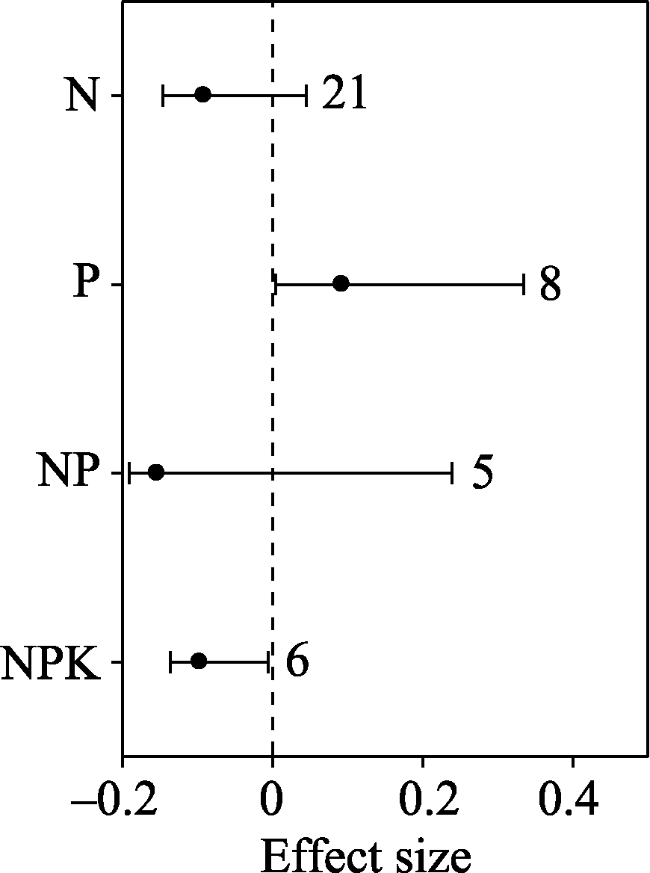
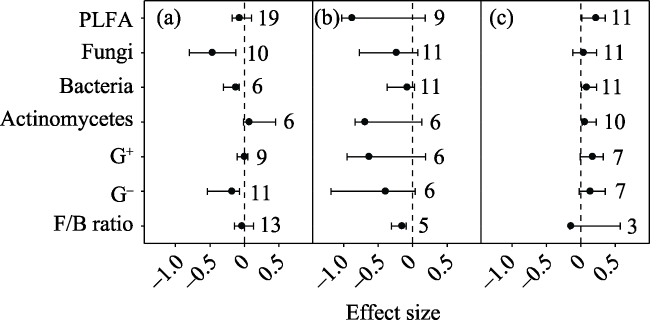
In order to investigate the general tendency of soil microbial community responses to fertilizers, a meta-analysis approach was used to synthesise observations on the effects of inorganic and organic fertilizer addition (N: nitrogen; P: phosphorus; NP: nitrogen and phosphorus; PK: phosphorus and potassium; NPK: nitrogen, phosphorus and potassium; OF: organic fertilizer; OF+NPK: organic fertilizer plus NPK) on soil microbial communities. Among the various studies, PK, NPK, OF and OF+NPK addition increased total phospholipid fatty acid (PLFA) by 52.0%, 19.5%, 334.3% and 58.3%, respectively; while NP, OF and OF+NPK addition increased fungi by 5.6%, 21.0% and 8.2%, respectively. NP, NPK and OF addition increased bacteria by 6.4%, 9.8% and 13.3%, respectively; while NP and NPK addition increased actinomycetes by 7.0% and 14.8%, respectively. Addition of ammonium nitrate rather than urea decreased gram-negative bacteria (G -). N addition increased total PLFA、bacteria and actinomycetes in croplands, but decreased fungi and bacteria in forests, and the F/B ratio in grasslands. NPK addition increased total PLFA in forests but not in croplands. The N addition rate was positively correlated with the effects of N addition on gram-positive bacteria (G +) and G -. Therefore, different fertilizers appear to have different effects on the soil microbial community. Organic fertilizers can have a greater positive effect on the soil microbial community than inorganic fertilizers. The effects of fertilizers on the soil microbial community varied with ecosystem types. The effect of N addition on the soil microbial community was related to both the forms of nitrogen that were added and the nitrogen addition rate.
Key words: ecosystem types; nitrogen addition rate; response ratio; soil PLFA
WANG Jiangwei , ZHANG Guangyu , YU Chengqun . A Meta-analysis of the Effects of Organic and Inorganic Fertilizers on the Soil Microbial Community[J]. Journal of Resources and Ecology, 2020 , 11(3) : 298 -303 . DOI: 10.5814/j.issn.1674-764X.2020.03.007
Fig. 1 Effect sizes of nitrogen (N), phosphorus (P), nitrogen+phosphorus (NP), phosphorus+potassium (PK), nitrogen+phosphorus+potassium (NPK), organic fertilizers (OF), and organic fertilizers + NPK (OF+NPK) on soil total PLFA. The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed line is drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
Fig. 2 Effect sizes of nitrogen (N), phosphorus (P), nitrogen+ phosphorus (NP), nitrogen+phosphorus+potassium (NPK), organic fertilizers (OF), and organic fertilizers+NPK (OF+ NPK) on soil (a) fungi, (b) bacteria and (c) actinomycetes. The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed lines are drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
Fig. 3 Effect sizes of nitrogen (N), phosphorus (P), nitrogen+phosphorus (NP), nitrogen+phosphorus+potassium (NPK) and organic fertilizers (OF) on soil (a) gram-positive bacteria, and (b) gram-negative bacteria. The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed lines are drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
Fig. 4 Effect sizes of nitrogen (N), phosphorus (P), nitrogen+phosphorus (NP), and nitrogen+phosphorus+potassium (NPK) on the ratio of soil fungi to bacteria (F/B ratio). The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed line is drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
Fig. 5 Effect sizes of nitrogen (N) on PLFA, fungi, bacteria, actinomycetes, gram-positive bacteria (G+) and gram-negative bacteria (G-) in (a) forest (b) grassland and (c) cropland. The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed lines are drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
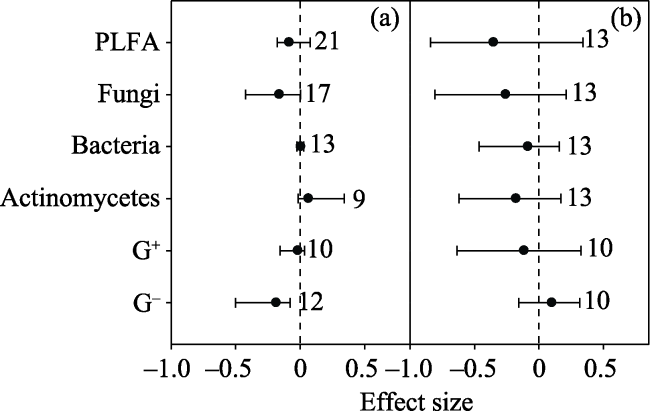
Fig. 6 Effect sizes of (a) ammonium nitrate and (b) urea on PLFA, fungi, bacteria, actinomycetes, gram-positive bacteria (G+) and gram-negative bacteria (G-). The error bars indicate effect sizes and 95% bootstrap confidence intervals. The effect was statistically significant if the 95%CI did not bracket zero. The dashed lines are drawn at effect size = 0. The sample size for each variable is shown next to the bar. |
Table 1 Relationships between the effect sizes of nitrogen addition on microbial indicators (total PLFA, fungi, bacteria, actinomycetes, G+, G-, and F/B ratio), and experimental variables (latitude, longitude, MAT, MAP, nitrogen addition rate, nitrogen load, and nitrogen addition duration) based on a random effects model with a continuous variable meta- analysis. |
| Variables | Slope | P | n | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Latitude | |||||||||||
| PLFA | 0.012 | 0.155 | 34 | ||||||||
| Fungi | 0.007 | 0.575 | 30 | ||||||||
| Bacteria | 0.006 | 0.477 | 26 | ||||||||
| G- | -0.003 | 0.812 | 22 | ||||||||
| Longitude | |||||||||||
| PLFA | -0.002 | 0.090 | 34 | ||||||||
| Fungi | -0.001 | 0.492 | 30 | ||||||||
| Bacteria | -0.001 | 0.466 | 26 | ||||||||
| G- | -0.0010 | 0.439 | 22 | ||||||||
| MAT | |||||||||||
| PLFA | 0.018 | 0.183 | 23 | ||||||||
| Fungi | 0.018 | 0.296 | 22 | ||||||||
| MAP | |||||||||||
| PLFA | 0.000 | 0.531 | 28 | ||||||||
| Fungi | 0.000 | 0.885 | 27 | ||||||||
| Bacteria | 0.000 | 0.907 | 23 | ||||||||
| G- | 0.000 | 0.361 | 21 | ||||||||
| Nitrogen addition rate | |||||||||||
| PLFA | 0.001 | 0.153 | 31 | ||||||||
| Fungi | 0.001 | 0.090 | 32 | ||||||||
| Bacteria | 0.001 | 0.051 | 28 | ||||||||
| Actinomycetes | 0.001 | 0.392 | 22 | ||||||||
| G+ | 0.002 | 0.015 | 22 | ||||||||
| G- | 0.002 | 0.001 | 24 | ||||||||
| F/B ratio | 0.000 | 0.513 | 21 | ||||||||
| Nitrogen load | |||||||||||
| Fungi | 0.000 | 0.425 | 21 | ||||||||
| Nitrogen duration | |||||||||||
| PLFA | -0.0001 | 0.906 | 30 | ||||||||
| Fungi | 0.0002 | 0.841 | 26 | ||||||||
| Bacteria | 0.0005 | 0.524 | 22 | ||||||||
| G- | 0.0006 | 0.434 | 21 | ||||||||
Note: G+ = gram-positive bacteria; G- = gram-negative bacteria; F/B ratio = ratio of fungi to bacteria; MAT = mean annual temperature; MAP = mean annual precipitation. |
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
Fu, G,
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}