
Journal of Resources and Ecology >
Species Abundance Distribution Patterns of a Toona ciliata Community in Xingdoushan Nature Reserve
Received date: 2019-03-11
Accepted date: 2019-05-31
Online published: 2019-10-11
Supported by
Public Welfare Research Project of Department of Science and Technology in Hubei Province(40 2012DBA40001)
Scientific Research Project of Department of Education in Hubei Province(B20160555)
Copyright
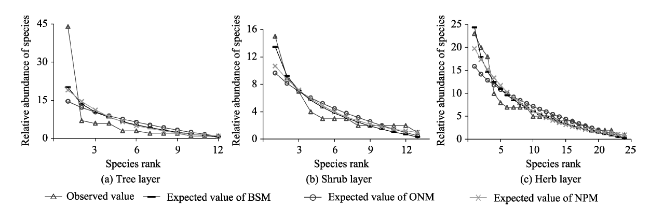
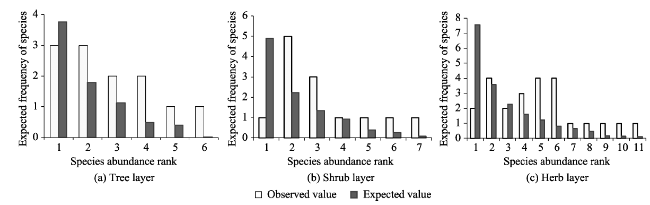
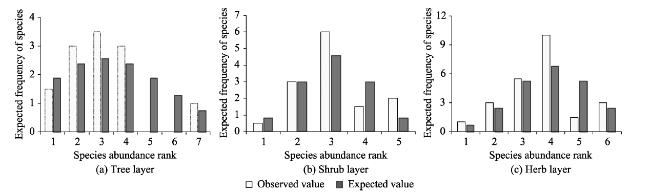
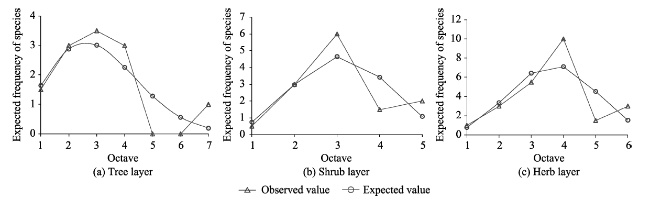
With the goal of model fitting species abundance distribution patterns of the tree, shrub and herb layers of the natural Toona ciliata community in Xingdoushan Nature Reserve, Enshi Autonomous Prefecture, Hubei Province, we used the data collected from the field survey and employed different ecological niche models. The models tested were the broken stick model (BSM), the overlapping niche model (ONM) and the niche preemption model (NPM), as well as three statistic models, the log-series distribution model (LSD), the log-normal distribution model (LND) and the Weibull distribution model (WDM). To determine the fitted model most suitable to each layer, the fitting effects were judged by criteria of the lowest value of Akaike Information Criterion (AIC), Chi-square and the K-S values with no significant difference (P>0.05) between the theoretical predictions and observed species abundance distribution values. The result showed: (1) The fitting suitability and goodness of fit of the tree, shrub and herb layers by using the three ecological niche models were ranked as: NPM>BSM>ONM. Of the three statistical models, by accepting the fitting results of the three layers, WDM was the best fitting model, followed by LND. By rejecting the fitting tests of the herb layer, LSD had the worst fitting effect. The goodness of the statistical models was ranked as: WDM>LND>LSD. In general, the statistical models had better fitting results than the ecological models. (2) T. ciliata was the dominant species of the tree layer. The species richness and diversity of the herb layer were much higher than those of either the tree layer or the shrub layer. The species richness and diversity of the shrub layer were slightly higher than those of the tree layer. The community evenness accorded to the following order: herb>shrub>tree. Considering the fitting results of the different layers, different ecological niche models or statistical models with optimal goodness of fit and ecological significance can be given priority to in studying the species abundance distribution patterns of T. ciliata communities.
WANG Yang , ZHU Shengjie , LI Jie , HE Xiuling , JIANG Xiongbo , ZHANG Min . Species Abundance Distribution Patterns of a Toona ciliata Community in Xingdoushan Nature Reserve[J]. Journal of Resources and Ecology, 2019 , 10(5) : 494 -503 . DOI: 10.5814/j.issn.1674-764X.2019.05.004
Table 1 Fitting test results of various ecological niche models |
| Model | Tree layer | Shrub layer | Herb layer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AIC value | χ2 test | K-S test | AIC value | χ2 test | K-S test | AIC value | χ2 test | K-S test | |||||||
| χ2 | P value | D | P value | χ2 | P value | D | P value | χ2 | P value | D | P value | ||||
| BSM | 30.679 | 39.461 | 0* | 0.333 | 0.518 | 2.461 | 6.516 | 0.837 | 0.308 | 0.570 | 21.342 | 9.426 | 0.991 | 0.167 | 0.893 |
| ONM | 29.911 | 73.339 | 0* | 0.417 | 0.249 | 10.258 | 7.820 | 0.729 | 0.308 | 0.570 | 31.309 | 13.637 | 0.914 | 0.208 | 0.675 |
| NPM | 33.077 | 43.877 | 0* | 0.250 | 0.847 | 5.713 | 4.245 | 0.962 | 0.231 | 0.879 | 27.675 | 7.405 | 0.998 | 0.167 | 0.893 |
P<0.01. |
Table 2 Parameters of LSD and LND and the fitting test results |
| Vegetation layer | LSD | LND | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| x value | α value | AIC value | χ2 test | K-S test | S0 value | λ value | AIC value | χ2 test | K-S test | |||||
| χ2 | P value | D | P value | χ2 | P value | D | P value | |||||||
| Tree layer | 0.952 | 3.959 | -0.469 | 0.999 | 0.318 | 0.500 | 0.441 | 2.566 | 0.280 | -8.530 | 3.989 | 0.408 | 0.429 | 0.541 |
| Shrub layer | 0.911 | 5.374 | 8.268 | 6.757 | 0.009** | 0.203 | 0.571 | 4.573 | 0.656 | 1.802 | 3.010 | 0.222 | 0.200 | 1.000 |
| Herb layer | 0.951 | 7.952 | 18.469 | 6.978 | 0.031* | 0.545 | 0.076 | 6.766 | 0.506 | 8.077 | 4.648 | 0.200 | 0.167 | 1.000 |
**P<0.01, *P<0.05. |
Table 3 Parameters of WDM and the fitting test results |
| Vegetation layer | a value | b value | c value | WDM | AIC value | χ2 test | K-S test | ||
|---|---|---|---|---|---|---|---|---|---|
| χ2 | P value | D | P value | ||||||
| Tree layer | 0 | 3.492 | 2.146 | $f(x)=0.147{{x}^{1.146}}{{e}^{\frac{-{{x}^{2.146}}}{14.636}}}$ | -5.516 | 5.598 | 0.133 | 0.286 | 0.938 |
| Shrub layer | 0 | 3.480 | 3.213 | $f(x)=0.059{{x}^{2.213}}{{e}^{\frac{-{{x}^{3.213}}}{54.9401}}}$ | 2.723 | 2.322 | 0.128 | 0.200 | 1.000 |
| Herb layer | 0 | 4.136 | 3.241 | $f(x)=0.033{{x}^{2.241}}{{e}^{\frac{-{{x}^{3.241}}}{99.626}}}$ | 8.254 | 4.832 | 0.089 | 0.333 | 0.893 |
Table 4 Correlation analysis of species diversity indices and parameters of the statistical models |
| parameter | x | α | S0 | λ | b | c | S | H | E |
|---|---|---|---|---|---|---|---|---|---|
| x | 1.000 | ||||||||
| α | 0.145 | 1.000 | |||||||
| S0 | 0.004 | 0.990 | 1.000 | ||||||
| λ | -0.815 | 0.456 | 0.576 | 1.000 | |||||
| b | 0.495 | 0.931 | 0.871 | 0.100 | 1.000 | ||||
| c | -0.499 | 0.785 | 0.864 | 0.909 | 0.506 | 1.000 | |||
| S | 0.414 | 0.961 | 0.912 | 0.190 | 0.996 | 0.582 | 1.000 | ||
| H | -0.031 | 0.984 | 0.999* | 0.605 | 0.853 | 0.882 | 0.897 | 1.000 | |
| E | -0.456 | 0.814 | 0.888 | 0.888 | 0.547 | 0.999* | 0.621 | 0.904 | 1.000 |
*P<0.05. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
The State Council of the People’s Republic of China. 1999. List of state key protected wild plants (the first directory). Plant Journal, (5):4-11.
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}