
Journal of Resources and Ecology >
Response of Microbial Communities in Soil to Multi-level Warming in a Highland Barley System of the Lhasa River
First author: FU Gang, E-mail: fugang@igsnrr.ac.cn
Received date: 2018-12-28
Accepted date: 2019-03-05
Online published: 2019-07-30
Supported by
National Natural Science Foundation of China (31370458, 31600432, 41807331)
Bingwei Outstanding Young Talents Program of Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences (2018RC202)
National Key Research Projects of China (2016YFC0502005, 2016YFC0502006, 2017YFA0604801)
Youth Innovation Research Team Project of Key Laboratory of Ecosystem Network Observation and Modeling (LENOM2016Q0002) and Tibet Science and Technology Major Projects of Pratacultural Industry (XZ201801NA02).
Copyright
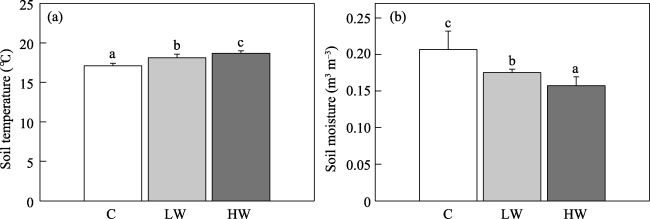
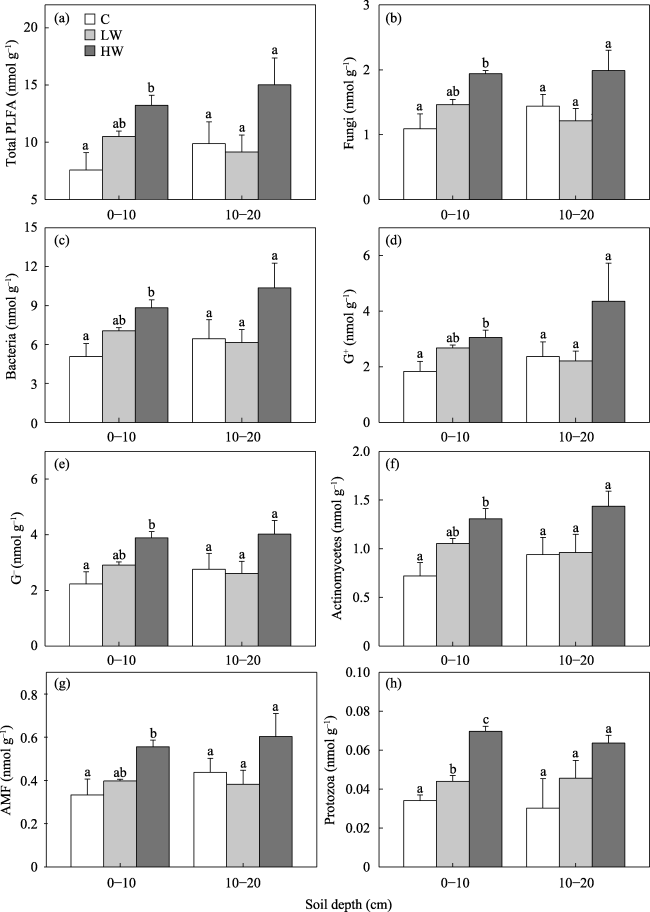
No studies have examined the effect of experimental warming on the microbial biomass and community composition of soil in agricultural ecosystem on the Qinghai-Tibet Plateau. Thus it is unclear whether the influences of experimental warming on microbial communities in soil are related to warming magnitude in croplands on this Plateau. This study performed warming experiment (control, low- and high-level) in a highland barley system of the Lhasa River in May 2014 to examine the correlation between the response of microbial communities in soil to warming and warming magnitude. Topsoil samples (0-10 and 10-20 cm) were collected on September 14, 2014. Experimental warming at both low and high levels significantly increased soil temperature by 1.02 ℃ and 1.59 ℃, respectively at the depth of 15 cm. Phospho lipid fatty acid (PLFA) method was used to determine the microbial community in soil. The low-level experimental warming did not significantly affect the soil’s total PLFA, fungi, bacteria, arbuscular mycorrhizal fungi (AMF), actinomycetes, gram-positive bacteria (G+), gram-negative bacteria (G-), protozoa, the ratio of fungi to bacteria (F/B ratio), and ratio of G+ to G- (G+/G- ratio) at the 0-10 and 10-20 cm depth. The low-level experimental warming also did not significantly alter the composition of microbial community in soil at the 0-10 and 10-20 cm depth. The high-level experimental warming significantly increased total PLFA by 74.4%, fungi by 78.0%, bacteria by 74.0%, AMF by 66.9%, actinomycetes by 81.4%, G+ by 67.0% and G- by 74.4% at the 0-10 cm depth rather than at 10-20 cm depth. The high-level experimental warming significantly altered microbial community composition in soil at the 0-10 cm depth rather than at 10-20 cm depth. Our findings suggest that the response of microbial communities in soil to warming varied with warming magnitudes in the highland barley system of the Lhasa River.
FU Gang , SUN Wei , LI Shaowei , ZHONG Zhiming . Response of Microbial Communities in Soil to Multi-level Warming in a Highland Barley System of the Lhasa River[J]. Journal of Resources and Ecology, 2019 , 10(4) : 373 -378 . DOI: 10.5814/j.issn.1674-764X.2019.04.004
Fig. 1 Warming effects on soil (a) temperature and (b) moisture |
Table 1 Two-way ANOVA for effects of warming (W) and soil depth (SD) on soil microbial biomass |
| Model | PLFA (nmol g-1) | Fungi (nmol g-1) | Bacteria(nmol g-1) | AMF(nmol g-1) | Actinomycetes(nmol g-1) | G+(nmol g-1) | G-(nmol g-1) | Protozoa(nmol g-1) | F/B | G+/G- | (i17:0+i15:0)/(a17:0+a15:0) | (cy17:0 w7c+cy19:0 w7c)/(16:1w7c+18:1w7c) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Warming (W) | 6.66* | 7.71** | 5.97* | 5.68* | 7.53** | 3.47 | 7.09** | 10.33** | 0.89 | 0.29 | 0.76 | 0.84 |
| Soil depth (SD) | 0.52 | 0.10 | 0.49 | 0.72 | 0.54 | 0.77 | 0.12 | 0.19 | 0.04 | 0.75 | 0.06 | 0.48 |
| W×SD | 0.80 | 1.17 | 0.68 | 0.41 | 0.63 | 0.95 | 0.48 | 0.13 | 1.01 | 1.37 | 1.44 | 1.37 |
Note: PLFA: phospho lipid fatty acid; AMF: arbuscular mycorrhizal fungi; G+: gram-positive bacteria; G-: gram-negative bacteria; F/B: the ratio of fungi to bacteria; G+/G-: the ratio of G+ to G-; *and ** mean P < 0.05 and P < 0.01, respectively. |
Fig. 2 Warming effects on (a) PLFA, (b) fungi, (c) bacteria, (d) G+, (e) G-, (f) actinomycetes, (g) AMF and (h) protozoa of soil. |
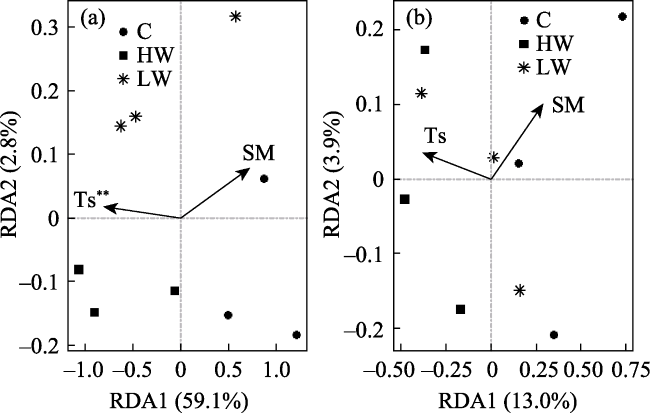
Fig. 3 Redundancy analysis (RDA) of PLFA at (a) 0-10 cm and (b) 10-20 cm |
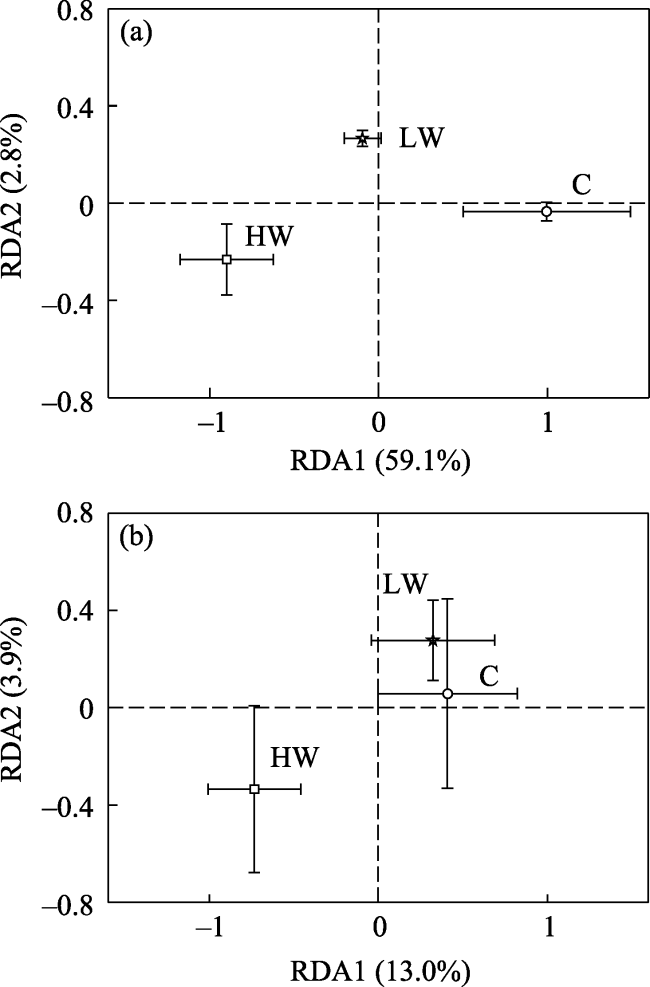
Fig. 4 Redundancy analysis (RDA) scores of PLFA at (a) 0-10 cm and (b) 10-20 cm |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
IPCC.2013.Summary for Policymakers. In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, United Kingdom and New York, NY, USA.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}