
Journal of Resources and Ecology >
Self-thinning Rules at Chinese Fir (Cunninghamia lanceolata) Plantations—Based on a Permanent Density Trial in Southern China
First author:Duan Aiguo, E-mail: duanag@163.com; FU Lihua, E-mail: 1308727478@qq.com
# The authors contributed equally to the work.
Received date: 2018-03-09
Accepted date: 2018-11-20
Online published: 2019-05-30
Supported by
The 12th and 13th Five-Year Plan of the National Scientific and Technological Support Projects (2015BAD09B01, 2016YFD0600302), Jiangxi Scientific and Technological innovation plan (201702) and National Natural Science Foundation of China (31570619, 31370629).
Copyright
Data selection and methods for fitting coefficients were considered to test the self-thinning law. The Chinese fir (Cunninghamia lanceolata) in even-aged pure stands with 26 years of observation data were applied to fit Reineke’s (1933) empirically derived stand density rule (N $propto$ d¯ -1.605, N = numbers of stems, d¯ = mean diameter), Yoda’s (1963) self-thinning law based on Euclidian geometry (v¯ $propto$ N -3/2, v¯ = tree volume), and West, Brown and Enquist’s (1997, 1999) (WBE) fractal geometry (w¯ $propto$ d¯ -8/3). OLS, RMA and SFF algorithms provided observed self-thinning exponents with the seven mortality rate intervals (2%-80%, 5%-80%, 10%-80%, 15%-80%, 20%-80%, 25%-80% and 30%-80%), which were tested against the exponents, and expected by the rules considered. Hope for a consistent allometry law that ignores species-specific morphologic allometric and scale differences faded. Exponents α of N $propto$ d¯α, were significantly different from -1.605 and -2, not expected by Euclidian fractal geometry; exponents β of w¯ $propto$ Nβ varied around Yoda’s self-thinning slope -3/2, but was significantly different from -4/3; exponent γ of w¯ $propto$ d¯γ tended to neither 8/3 nor 3.
Key words: Chinese fir; self-thinning; stand density; mortality rate
DUAN Aiguo , FU Lihua , ZHANG Jianguo . Self-thinning Rules at Chinese Fir (Cunninghamia lanceolata) Plantations—Based on a Permanent Density Trial in Southern China[J]. Journal of Resources and Ecology, 2019 , 10(3) : 315 -323 . DOI: 10.5814/j.issn.1674-764X.2019.03.010
Table 1 Summary of the stand attributes of the Chinese fir stands |
| Stand attribute | Mean | S.D.a | Minimum | Maximum |
|---|---|---|---|---|
| Age (years) | 16 | 6 | 2 | 26 |
| Density (stems ha-1) | 5783 | 2216 | 2516 | 10000 |
| Diameter at breast height (cm) | 10.98 | 4.02 | 6.23 | 11.24 |
| Average stem volume (dm3) | 0.07 | 0.05 | 0.01 | 0.32 |
a S.D.: Standard Deviation. |
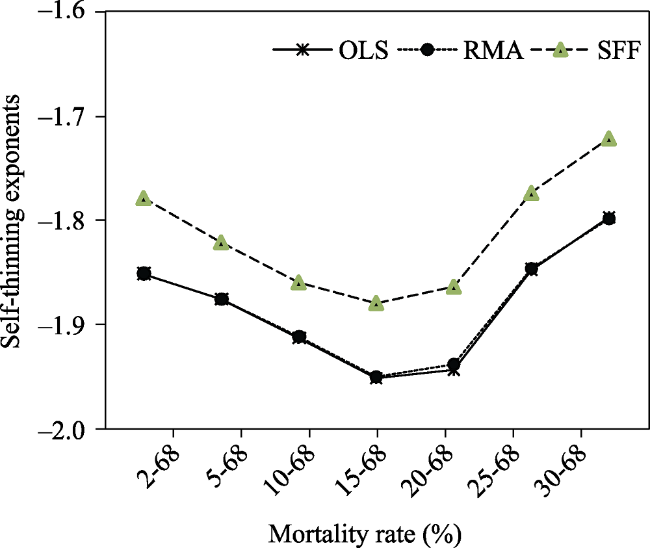
Table 2 Results of the Reineke’s slopes of the self-thinning line for the LnN versus Ln $\overline{d}$ relationships in Fujian plots |
| Mortality rate (%) | Sample size | Regression method | R2 a | Estimated slopes | S.E. b | 95% CI c |
|---|---|---|---|---|---|---|
| 2-68 | 66 | OLS | 0.926 | -1.851 | 0.063 | -1.977, -1.725 |
| RMA | 0.926 | -1.851 | 0.063 | -1.946, -1.756 | ||
| SFF | 0.932 | -1.778 | 0.064 | -1.902, -1.653 | ||
| 5-68 | 59 | OLS | 0.943 | -1.876 | 0.059 | -1.995, -1.757 |
| RMA | 0.944 | -1.876 | 0.050 | -1.972, -1.780 | ||
| SFF | 0.957 | -1.821 | 0.060 | -1.938, -1.705 | ||
| 10-68 | 51 | OLS | 0.945 | -1.913 | 0.064 | -2.043, -1.784 |
| RMA | 0.945 | -1.912 | 0.061 | -2.036, -1.797 | ||
| SFF | 0.947 | -1.860 | 0.062 | -1.981, -1.738 | ||
| 15-68 | 43 | OLS | 0.928 | -1.952 | 0.082 | -2.118, -1.787 |
| RMA | 0.928 | -1.951 | 0.079 | -2.113, -1.810 | ||
| SFF | 0.975 | -1.880 | 0.084 | -2.045, -1.716 | ||
| 20-68 | 37 | OLS | 0.919 | -1.944 | 0.093 | -2.134, -1.755 |
| RMA | 0.920 | -1.939 | 0.091 | -2.134, -1.790 | ||
| SFF | 0.934 | -1.864 | 0.093 | -2.046, -1.682 | ||
| 25-68 | 28 | OLS | 0.921 | -1.847 | 0.102 | -2.055, -1.638 |
| RMA | 0.920 | -1.846 | 0.118 | -2.08, -1.634 | ||
| SFF | 0.935 | -1.773 | 0.090 | -1.950, -1.595 | ||
| 30-68 | 25 | OLS | 0.916 | -1.797 | 0.109 | -2.022, -1.572 |
| RMA | 0.913 | -1.798 | 0.132 | -2.039, -1.549 | ||
| SFF | 0.928 | -1.720 | 0.097 | -1.909, -1.530 |
a R is correlation coefficient. b S.E. is the standard errors associated with the slope. c 95% confidence intervals. |
Fig. 1 The Reineke’s self-thinning exponents with OLS, RMA and SFF change across the seven mortality rate classes. |
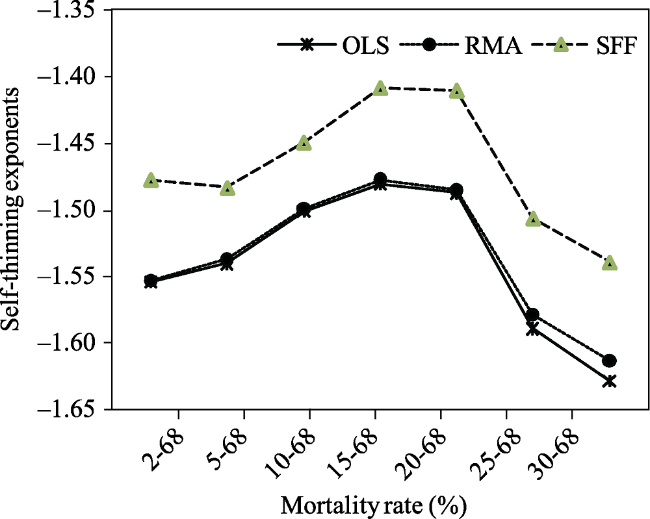
Table 3 Results of the Yoda’s slopes of the self-thinning line for the Lnw versus LnN relationships in Fujian plots |
| Mortality rate (%) | Sample size | Regression method | R2 a | Estimated slopes | S.E. b | 95% CI c |
|---|---|---|---|---|---|---|
| 2-68 | 66 | OLS | 0.895 | -1.554 | 0.063 | -1.680, -1.428 |
| RMA | 0.897 | -1.553 | 0.046 | -1.637, -1.467 | ||
| SFF | 0.913 | -1.477 | 0.063 | -1.601, -1.353 | ||
| 5-68 | 59 | OLS | 0.928 | -1.540 | 0.055 | -1.649, -1.430 |
| RMA | 0.929 | -1.537 | 0.046 | -1.633, -1.459 | ||
| SFF | 0.951 | -1.483 | 0.052 | -1.585, -1.381 | ||
| 10-68 | 51 | OLS | 0.931 | -1.501 | 0.056 | -1.615, -1.388 |
| RMA | 0.932 | -1.499 | 0.050 | -1.607, -1.412 | ||
| SFF | 0.944 | -1.449 | 0.054 | -1.554, -1.343 | ||
| 15-68 | 43 | OLS | 0.904 | -1.480 | 0.071 | -1.625, -1.336 |
| RMA | 0.906 | -1.477 | 0.061 | -1.621, -1.368 | ||
| SFF | 0.915 | -1.408 | 0.069 | -1.543, -1.272 | ||
| 20-68 | 37 | OLS | 0.899 | -1.487 | 0.080 | -1.649, -1.325 |
| RMA | 0.911 | -1.485 | 0.067 | -1.628, -1.359 | ||
| SFF | 0.926 | -1.410 | 0.063 | -1.534, -1.286 | ||
| 25-68 | 28 | OLS | 0.898 | -1.589 | 0.099 | -1.793, -1.384 |
| RMA | 0.898 | -1.578 | 0.102 | -1.821, -1.434 | ||
| SFF | 0.915 | -1.506 | 0.093 | -1.688, -1.324 | ||
| 30-68 | 25 | OLS | 0.895 | -1.628 | 0.110 | -1.856, -1.400 |
| RMA | 0.893 | -1.613 | 0.121 | -1.883, -1.443 | ||
| SFF | 0.904 | -1.539 | 0.102 | -1.738, -1.340 |
Fig. 2 The Yoda’s self-thinning exponents with OLS, RMA and SFF change across the seven mortality rate classes. |
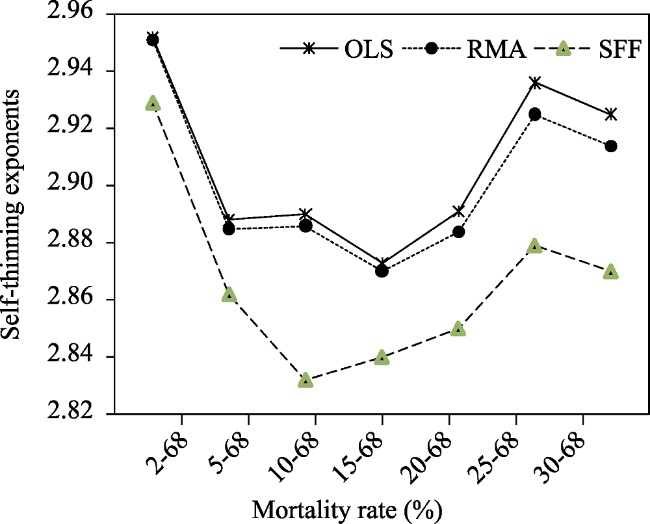
Table 4 Results of the WBE slopes of the self-thinning line for the Ln$w$ versus LnN relationships in Fujian plots |
| Mortality rate (%) | Sample size | Regression method | R2 a | Estimated slopes | S.E. b | 95% CI c |
|---|---|---|---|---|---|---|
| 2-68 | 83 | OLS | 0.984 | 2.952 | 0.041 | 2.871, 3.034 |
| RMA | 0.985 | 2.951 | 0.032 | 2.891, 3.022 | ||
| SFF | 0.991 | 2.929 | 0.041 | 2.849, 3.008 | ||
| 5-68 | 59 | OLS | 0.982 | 2.888 | 0.051 | 2.785, 2.99 |
| RMA | 0.982 | 2.885 | 0.035 | 2.827, 2.963 | ||
| SFF | 0.987 | 2.862 | 0.049 | 2.766, 2.957 | ||
| 10-68 | 51 | OLS | 0.978 | 2.873 | 0.061 | 2.749, 2.996 |
| RMA | 0.978 | 2.886 | 0.041 | 2.805, 2.968 | ||
| SFF | 0.974 | 2.832 | 0.058 | 2.727, 2.953 | ||
| 15-68 | 43 | OLS | 0.970 | 2.890 | 0.078 | 2.732, 3.048 |
| RMA | 0.971 | 2.87 | 0.063 | 2.772, 3.019 | ||
| SFF | 0.978 | 2.840 | 0.065 | 2.705, 2.958 | ||
| 20-68 | 37 | OLS | 0.972 | 2.891 | 0.082 | 2.725, 3.057 |
| RMA | 0.972 | 2.884 | 0.077 | 2.761, 3.054 | ||
| SFF | 0.983 | 2.850 | 0.081 | 2.705, 2.958 | ||
| 25-68 | 28 | OLS | 0.961 | 2.936 | 0.111 | 2.708, 3.163 |
| RMA | 0.962 | 2.925 | 0.109 | 2.708, 3.163 | ||
| SFF | 0.953 | 2.879 | 0.106 | 2.692, 3.009 | ||
| 30-68 | 25 | OLS | 0.963 | 2.925 | 0.117 | 2.682, 3.167 |
| RMA | 0.963 | 2.914 | 0.115 | 2.727, 3.164 | ||
| SFF | 0.978 | 2.870 | 0.100 | 2.671, 3.087 |
Fig. 3 The WBE’s self-thinning exponents with OLS, RMA and SFF change across the seven mortality rate classes. |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}