
Journal of Resources and Ecology >
Temperature Affects New Carbon Input Utilization by Soil Microbes: Evidence based on a Rapid δ13C Measurement Technology
Received date: 2018-12-10
Accepted date: 2019-01-30
Online published: 2019-03-30
Supported by
National Key Research and Development Program of China (2016YFA0600104, 2016YFC0500102)
Natural Science Foundation of China (31770655, 41671045)
and Program of Youth Innovation Research Team Project (LENOM2016Q0005).
Copyright
Strong and rapid responses of soil microbial respiration to pulses, such as those from available soil organic matter (SOM) or water input from precipitation (especially in arid areas), are common. However, how soil microbes utilize new SOM inputs and the effects that temperature may have on their activities are unclear owing to the limitation in the application of traditional isotopic techniques at minute scales. In the present study, we developed a system of measuring 12CO2 and δ13C minutely and synchronously under controlled incubation temperatures, i.e., for 48 h at 7, 10, 15, 20, and 25 °C, to explore the carbon utilization strategies of soil microbes. We measured the respiration rates of soil microbes in response to different carbon sources, i.e., added glucose (Rg) and initial SOM (Rs), as well as the total respiration rate (Rt). All responses were rapid and characterized by unimodal curves. Furthermore, the characteristic values of these curves, such as the maximum of rate (R-max), the time required to achieve R-max, and the ratio of the duration of R-max to that of 1/2 R-max, were all dependent on incubation temperature. Interestingly, temperature greatly influenced the strategy that microorganisms employed to utilize different carbon sources. The effects of temperature on the intensity of the microbial respiratory response and the ratio of Rg/Rs are important for evaluating the effect of land-use changes or variations in seasonal temperature on SOM turnover and should be considered in ecological models in future studies.
Key words: soil respiration; decomposition; isotopic; pulse; soil organic matter; turnover
CAO Yingqiu , ZHANG Zhen , XU Li , CHEN Zhi , HE Nianpeng . Temperature Affects New Carbon Input Utilization by Soil Microbes: Evidence based on a Rapid δ13C Measurement Technology[J]. Journal of Resources and Ecology, 2019 , 10(2) : 202 -212 . DOI: 10.5814/j.issn.1674-764X.2019.02.011
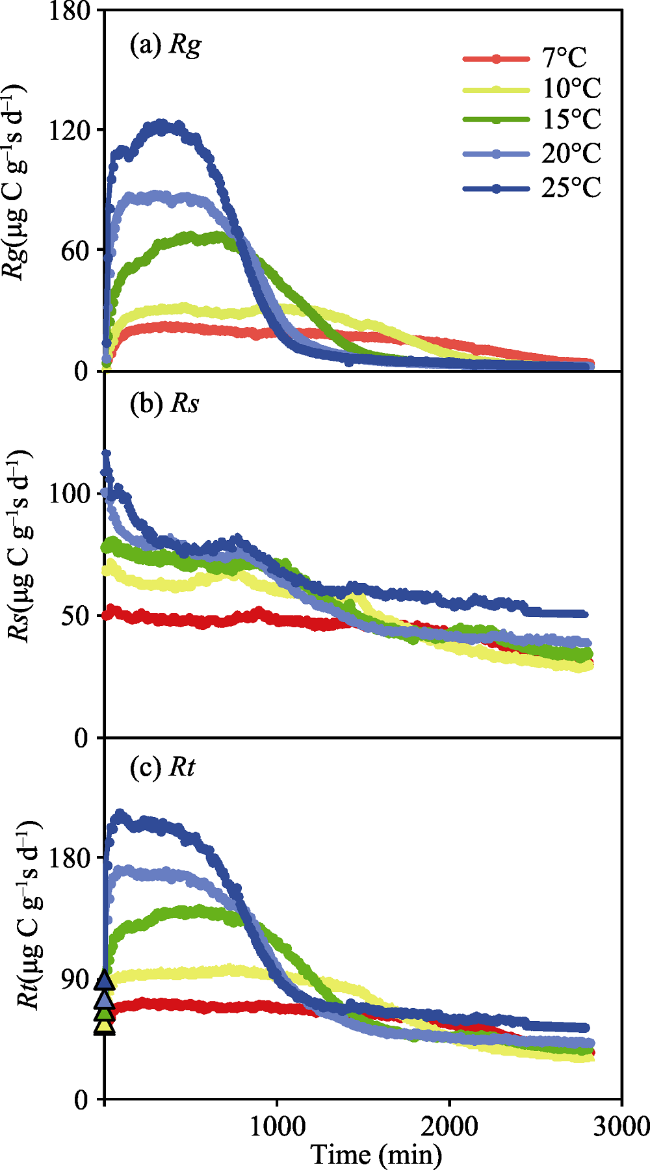
Fig. 1 Changes in the respiration rate using glucose (Rg, (a)) and soil organic matter (Rs, (b)), and the total respiration rate (Rt, (c)) in a glucose addition treatment conducted at a minute scale. |
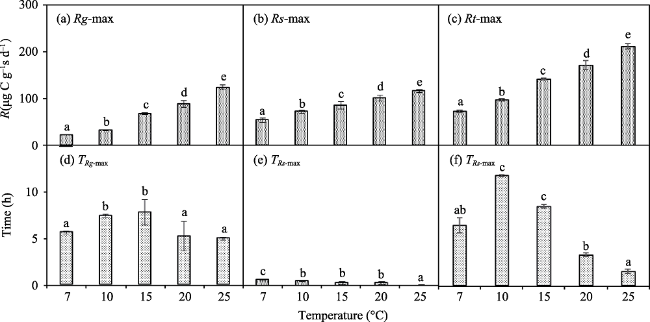
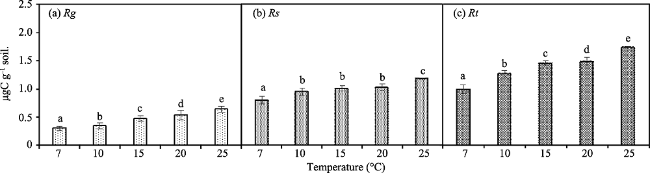
Fig. 2 The Rg-max, Rs-max, Rt-max, TRg-max, TRs-max, and TRt-max at different temperatures. |
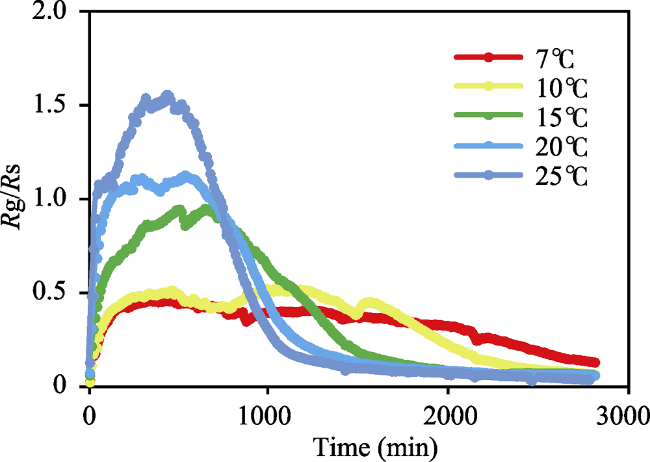
Fig. 3 The changing trend for the ratio of Rg/Rs at different incubation temperatures |
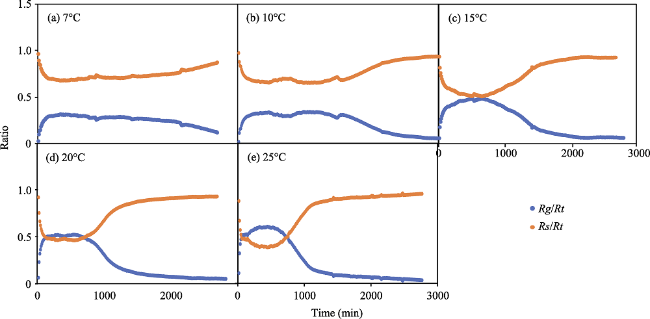
Fig. 4 Changes in the ratio of Rg/Rt and Rs/Rt at different incubation temperatures |
Fig. 5 The accumulation of C emission from Rg, Rs, and Rt within 48 h at different temperatures. |
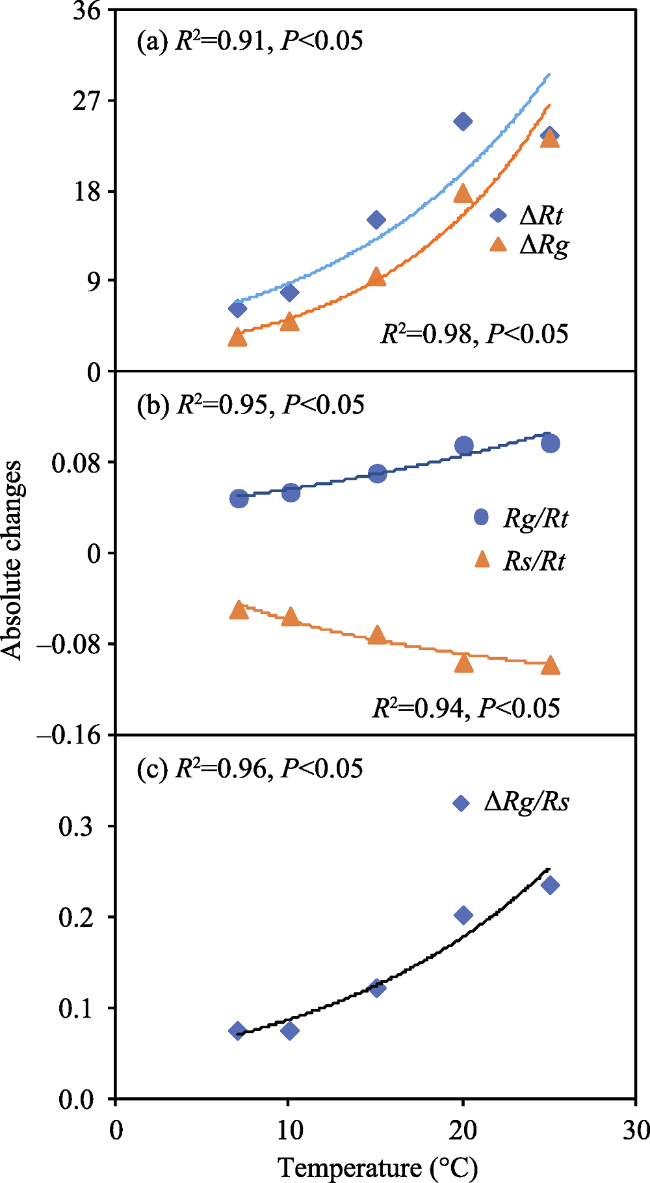
Fig. 6 The relationship of the absolute changes in Rt (ΔRt), Rg (ΔRg), Rg/Rt (ΔRg/Rt), Rs/Rt (ΔRs/Rt), and Rg/Rs (ΔRg/Rs) with temperature under adequate substrate supply. |
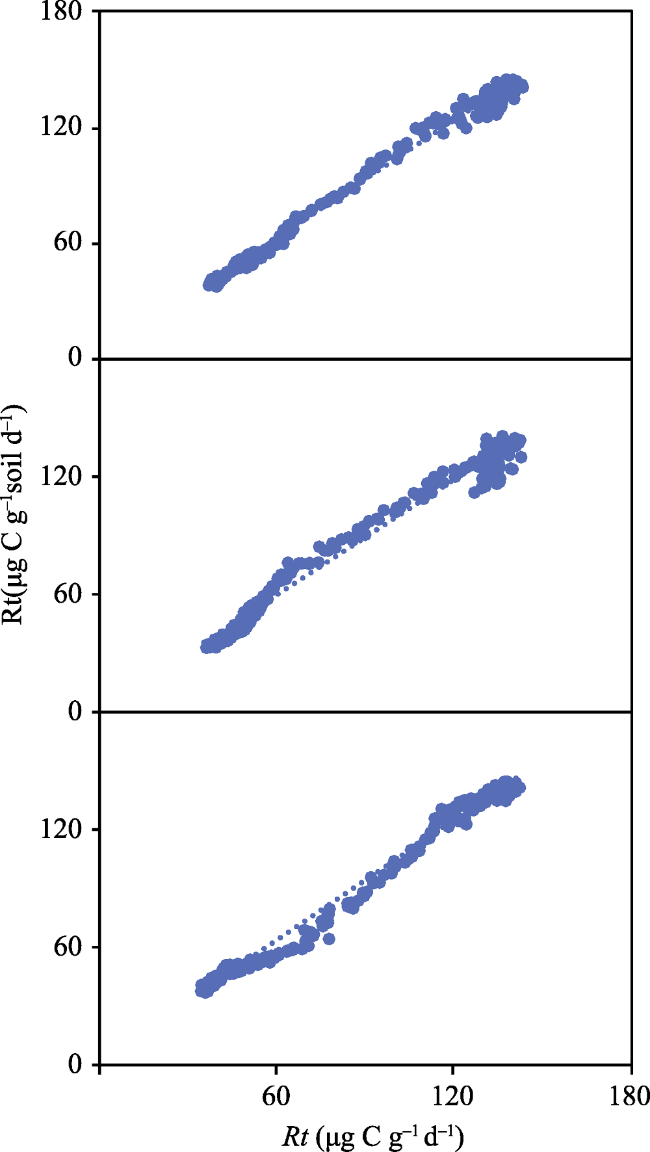
Fig. 7 Correlation test of three groups of repeated data |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}