
Journal of Resources and Ecology >
Soil Respiration Dynamics and Influencing Factors in Typical Steppe of Inner Mongolia under Long-term Nitrogen Addition
Received date: 2018-10-17
Accepted date: 2018-12-29
Online published: 2019-03-30
Supported by
National Key Research and Development Program of China (2017YFC0503805)
National Natural Science Foundation of China (31770519).
Copyright
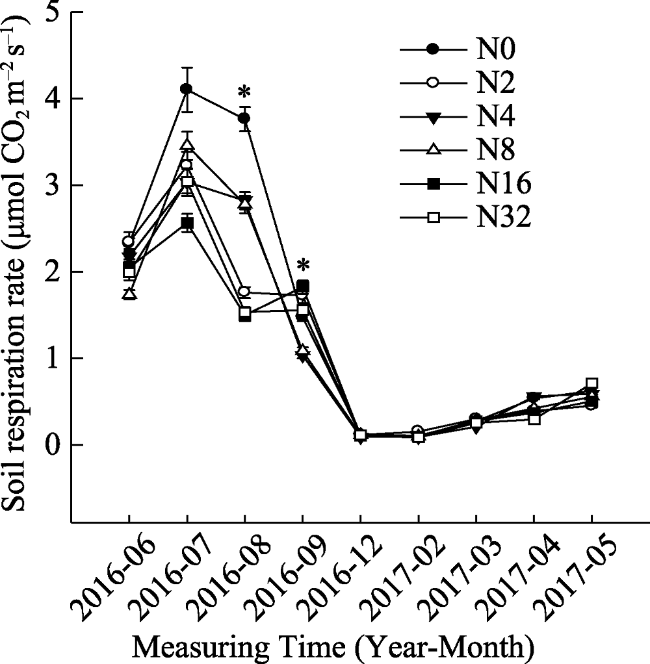
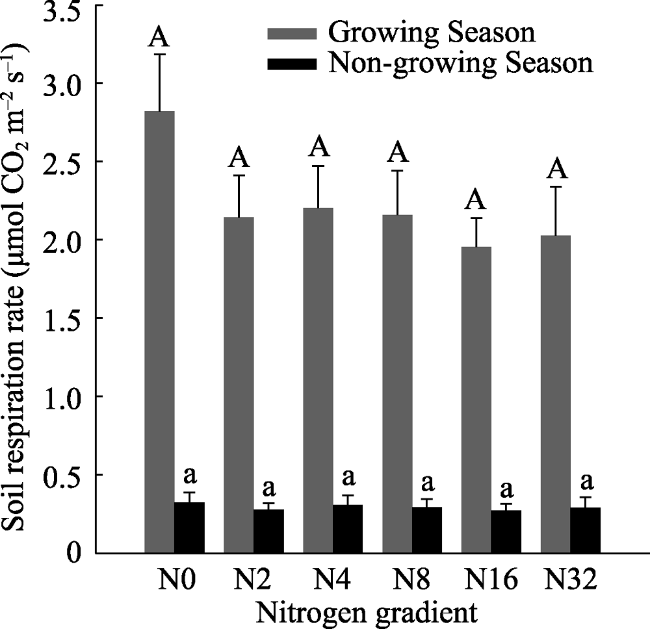
We investigated soil respiration (Rs) dynamics and influencing factors under different nitrogen (N) addition levels (0, 2, 4, 8, 16, 32 g m-2 yr-1) on typical grassland plots in Inner Mongolia. We measured soil respiration, temperature, moisture and nutrients. We found that N addition did not change dynamic characteristics of Rs; daily and seasonal dynamics followed a single peak curve. N addition reduced Rs during the growing season. Rs under N2, N4, N8, N16 and N32 treatments decreased by 24.00%, 21.93%, 23.49%, 30.78% and 28.20% in the growing season, respectively, compared to the N0 treatment. However, Rs in the non-growing season was not different across treatments. Rs was significantly positively correlated with soil temperature and moisture and these two factors accounted for 72%-97% and 74%-82% of variation in Rs, respectively. The soil respiration temperature sensitivity (Q10) was between 2.27 and 4.16 and N addition reduced Q10 except in the N8 treatment.
DU Wei , WU Shanmei , NIE Cheng , LI Yue , SHAO Rui , LIU Yinghui , SUN Nan . Soil Respiration Dynamics and Influencing Factors in Typical Steppe of Inner Mongolia under Long-term Nitrogen Addition[J]. Journal of Resources and Ecology, 2019 , 10(2) : 155 -162 . DOI: 10.5814/j.issn.1674-764X.2019.02.006
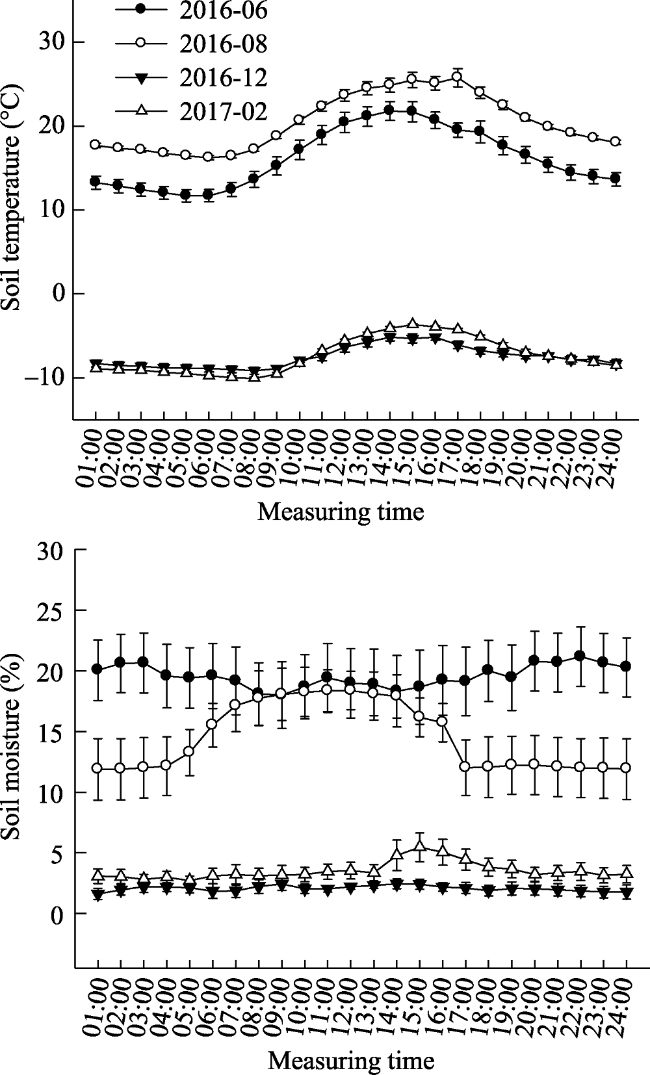
Fig. 1 Daily dynamic curve of soil temperature and moisture in different months (mean ± SE) |
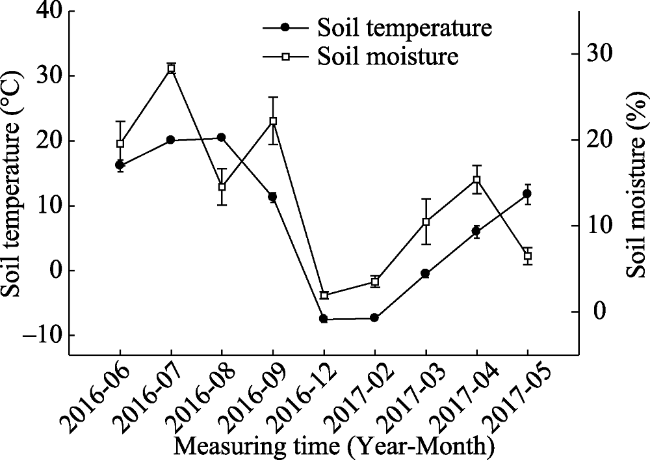
Fig. 2 Seasonal dynamic curve of soil temperature and moisture (mean ± SE) |
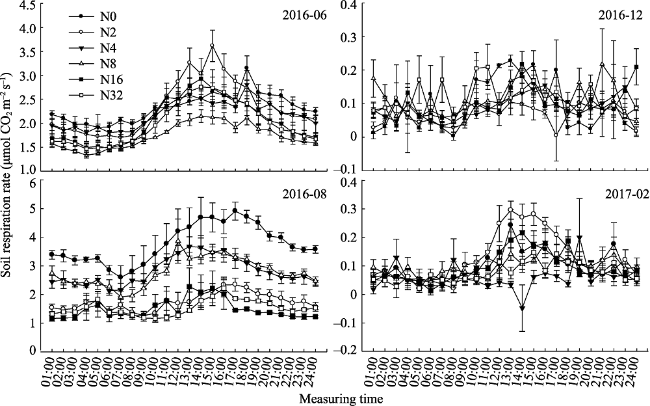
Fig. 3 Daily soil respiration rate in different months under different nitrogen additions (mean ± SE) |
Fig. 4 Seasonal dynamic curve of soil respiration rate (mean ± SE) |
Fig. 5 Dynamic soil respiration rate during the growing and non- growing seasons under different N addition levels (mean ± SE) |
Table 1 Function fitting of soil respiration (Rs) and soil temperature and moisture |
| N addition (g m-2 yr-1) | Rs and soil temperature | Rs and soil moisture | |||||
|---|---|---|---|---|---|---|---|
| Formula | R2 | P | Q10 | Formula | R2 | P | |
| 0 | Rs = 0.2245e0.1417T | 0.97 | <0.01 | 4.13 | Rs = 0.014M1.646 | 0.76 | <0.01 |
| 2 | Rs = 0.3851e0.0945 T | 0.75 | <0.05 | 2.57 | Rs = 0.017M1.499 | 0.80 | <0.01 |
| 4 | Rs = 0.2385e0.1250 T | 0.96 | <0.01 | 3.49 | Rs = 0.013M1.600 | 0.75 | <0.01 |
| 8 | Rs = 0.1736e0.1425 T | 0.95 | <0.01 | 4.16 | Rs = 0.015M1.535 | 0.75 | <0.01 |
| 16 | Rs = 0.4146e0.0819 T | 0.72 | <0.05 | 2.27 | Rs = 0.013M1.560 | 0.82 | <0.01 |
| 32 | Rs = 0.3644e0.0927 T | 0.77 | <0.05 | 2.53 | Rs = 0.014M1.530 | 0.74 | <0.01 |
Table 2 Pearson correlation between soil nutrients and soil respiration (Rs) |
| DOC | MBC | NH4+-N | NO3--N | TON | pH | |
|---|---|---|---|---|---|---|
| Rs (August 2016) | -0.079 | 0.604** | -0.582* | -0.483* | 0.088 | 0.615** |
| Rs (December 2016) | 0.378 | -0.076 | 0.390 | 0.149 | -0.226 | -0.154 |
Note: * P<0.05,** P<0.01 |
Table 3 Pearson correlation between nitrogen (N) addition and soil nutrients |
| N addition (August 2016) | N addition (December 2016) | |
|---|---|---|
| DOC | 0.207 | 0.613** |
| MBC | -0.676** | -0.841*** |
| NH4+-N | 0.700** | 0.753*** |
| NO3--N | 0.743*** | 0.656** |
| TON | -0.011 | -0.335 |
| pH | -0.962*** | -0.951*** |
Note: ** P<0.01, *** P<0.001 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
Haszulah.2015. Effects of nitrogen addition to soil respiration in Stipa krylovii steppe. Hohhot, Inner Mongolia Normal University. (in Chinese)
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}