
Journal of Resources and Ecology >
Soil Priming Effect Mediated by Nitrogen Fertilization Gradients in a Semi-arid Grassland, China
Received date: 2018-10-17
Accepted date: 2018-12-20
Online published: 2019-03-30
Supported by
National Natural Science Foundation of China (31770519)
National Key Research and Development Program of China (2017YFC0503805).
Copyright
The priming effect is well acknowledged in soil systems but the effect of nitrogen (N) fertilization remains elusive. To explore how N modifies the priming effect in soil organic matter (SOM), one in situ experiment with 13C labeled glucose addition (0.4 mg C g-1 soil, 3.4 atom % 13C) was conducted on soil plots fertilized with three gradients of urea (0, 4 and 16 g N m-2 yr-1). After glucose addition, the soil CO2 concentration and phospholipid fatty acid (PLFA) were measured on day 3, 7, 21 and 35. The study found that N fertilization decreased soil CO2, PLFA and the fungi to bacteria ratio. Glucose triggered the strongest positive priming in soil at 0 g N m-2 yr-2, meanwhile N fertilization decreased SOM-derived CO2. Soil at 4 g N m-2 yr-2 released the largest amount of glucose-derived carbon (C), likely due to favorable nutrient stoichiometry between C and N. Stable microbial community biomass and composition during early sampling suggests “apparent priming” in this grassland. This study concludes that N fertilization inhibited soil priming in semi-arid grassland, and shifted microbial utilization of C substrate from SOM to added labile C. Diverse microbial functions might be playing a crucial role in soil priming and requires attention in future N fertilization studies.
LI Yue , NIE Cheng , SHAO Rui , DU Wei , LIU Yinghui . Soil Priming Effect Mediated by Nitrogen Fertilization Gradients in a Semi-arid Grassland, China[J]. Journal of Resources and Ecology, 2019 , 10(2) : 147 -154 . DOI: 10.5814/j.issn.1674-764X.2019.02.005
Table 1 Repeated-measures ANOVA for soil CO2, total PLFA, bacteria, fungi and the F:B ratio among treatments in four sampling days. |
| Source of variation | Soil CO2 | Total PLFA | Bacteria | Fungi | F:B ratio |
|---|---|---|---|---|---|
| Day | 1.13 | 44.53*** | 40.16*** | 28.55*** | 0.41 |
| Nitrogen | 8.21** | 23.43*** | 0.73 | 22.24*** | 19.77*** |
| Glucose | 134.83*** | 142.39*** | 0.01 | 8.57** | 6.58* |
| Day × Nitrogen | 1.66 | 1.17 | 7.87*** | 2.94** | 0.85 |
| Day × Glucose | 20.01*** | 19.16*** | 63.38*** | 15.93*** | 14.19*** |
| Nitrogen × Glucose | 1.90 | 2.01 | 1.58 | 0.01 | 0.02 |
| Day × Nitrogen ×Glucose | 1.94 | 1.86 | 7.00*** | 2.37 | 0.38 |
Note: * indicates significant difference at P < 0.05, ** indicates significant difference at P < 0.01, *** indicates significant difference at P < 0.001. |
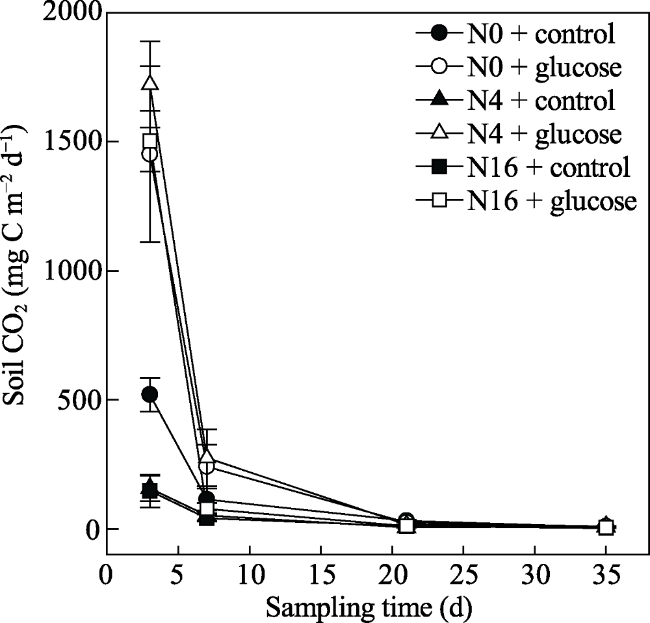
Fig. 1 Dynamics of soil CO2 among treatments in four sampling days |
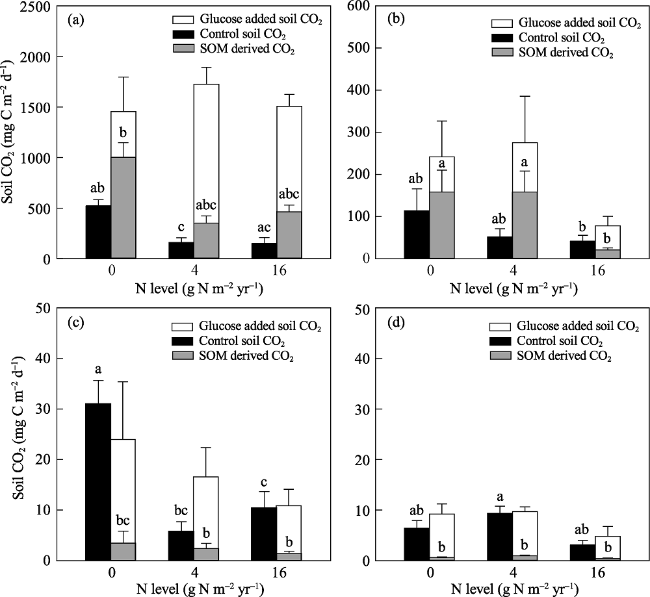
Fig. 2 Soil CO2 in the control treatment and soil CO2 and soil organic matter derived CO2 in the 13C labeled glucose treatment on day 3(a), 7(b), 21(c), and 35(d). |
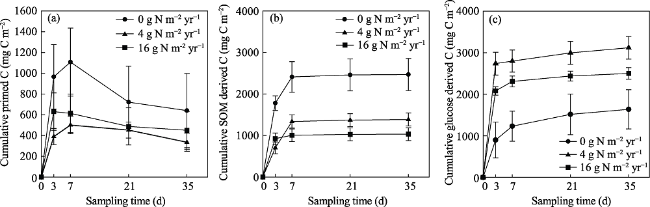
Fig. 3 Cumulative primed carbon (a), soil organic matter derived carbon (b) and glucose-derived carbon (c) in four sampling days |
Table 2 Spearman correlations between N levels and total PLFA, bacteria, fungi and the F:B ratio averaged by sampling days. |
| Treatment | Variable | Coefficient |
|---|---|---|
| Control | Total PLFA | -0.621* |
| Bacteria | -0.443 | |
| Fungi | -0.857** | |
| F:B ratio | -0.828** | |
| Glucose | Total PLFA | -0.680* |
| Bacteria | 0.266 | |
| Fungi | -0.710** | |
| F:B ratio | -0.917** |
Note: * indicates significant difference at P < 0.05, ** indicates significant difference at P < 0.01. |
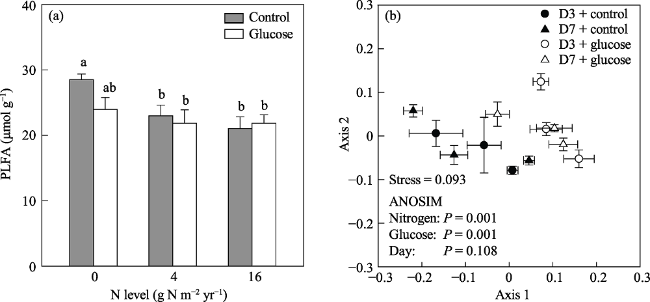
Fig. 4 Mean PLFA biomass on day 3 and 7 (a) and non-metric multidimensional scaling ordination of PLFA biomarkers (b) among treatments on day 3 and 7 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}