
Journal of Resources and Ecology >
Remote Sensing Indices to Measure the Seasonal Dynamics of Photosynthesis in a Southern China Subtropical Evergreen Forest
Received date: 2018-10-11
Accepted date: 2018-11-12
Online published: 2019-03-30
Supported by
National Key Research and Development Program of China (2017YFC0503803)
National Natural Science Foundation of China (41571192)
Natural Science Foundation of Hebei, China (D2016302002)
Science and Technology Planning Project of Hebei, China (17390313D).
Copyright
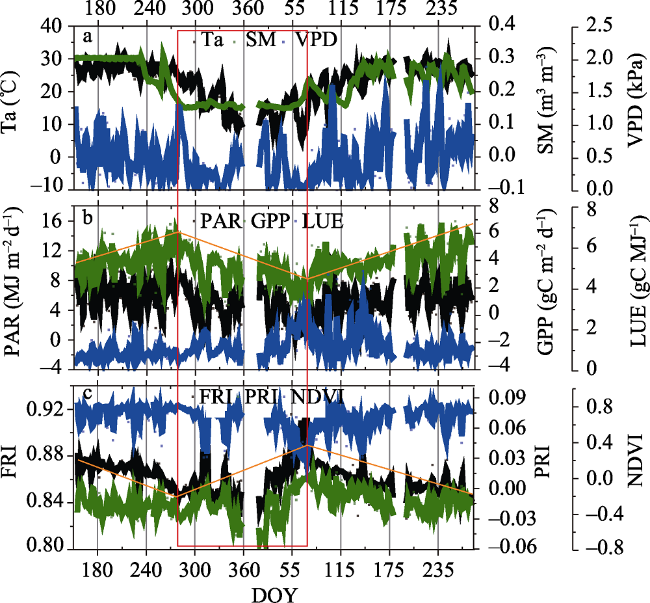
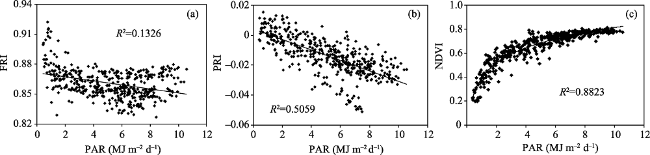
The accurate measurement of the dynamics of photosynthesis in China’s subtropical evergreen forest ecosystems is an important contribution to carbon (C) sink estimates in global terrestrial ecosystems and their responses to climate change. Eddy covariance has historically been the only direct method to assess C flux of whole ecosystems with high temporal resolution, but it suffers from limited spatial resolution. During the last decade, continuous global monitoring of plant primary productivity from spectroradiometer sensors on flux towers and satellites has extended the temporal and spatial coverage of C flux observations. In this study, we evaluated the performance of two physiological remote sensing indices, fluorescence reflectance index (FRI) and photochemical reflectance index (PRI), to measure the seasonal variations of photosynthesis in a subtropical evergreen forest ecosystem using continuous canopy spectral and flux measurements in the Dinghushan Nature Reserve in southern China. The more commonly used NDVI has been shown to be saturated and mainly affected by illumination (R2=0.88, p < 0.001), but FRI and PRI could better track the seasonal dynamics of plant photosynthetic functioning by comparison and are less affected by illumination (R2=0.13 and R2=0.51, respectively) at the seasonal scale. FRI correlated better with daily gross primary production (GPP) in the morning hours than in the afternoon hours, in contrast to PRI which correlated better with light-use efficiency (LUE) in the afternoon hours. Both FRI and PRI could show greater correlations with GPP and LUE respectively in the senescence season than in the recovery-growth season. When incident PAR was taken into account, the relationship between GPP and FRI was improved and the correlation coefficient increased from 0.22 to 0.69 (p < 0.001). The strength of the correlation increased significantly in the senescence season (R 2=0.79, p < 0.001). Our results demonstrate the application of FRI and PRI as physiological indices for the accurate measurement of the seasonal dynamics of plant community photosynthesis in a subtropical evergreen forest, and suggest these indices may be applied to carbon cycle models to improve the estimation of regional carbon budgets.
SUN Leigang , WANG Shaoqiang , Robert A. MICKLER , CHEN Jinghua , YU Quanzhou , QIAN Zhaohui , ZHOU Guoyi , MENG Ze . Remote Sensing Indices to Measure the Seasonal Dynamics of Photosynthesis in a Southern China Subtropical Evergreen Forest[J]. Journal of Resources and Ecology, 2019 , 10(2) : 112 -126 . DOI: 10.5814/j.issn.1674-764X.2019.02.002
Fig. 1 Seasonal variation of climatic factors, GPP, LUE and remote sensing indices |
Fig. 2 Relationships of daily climatic factors (PAR, Ta and VPD) with GPP (a-c) and LUE (d-f) from 1 June 2014 to 30 September 2015 |
Fig. 3 Relationships of PAR with FRI (a), PRI (b), and NDVI (c) from 1 June 2014 to 30 September 2015 |
Fig. 4 Seasonal variation of GPP and various FRI |
Fig. 5 Relationships of daily GPP with FRI (a), FRI_am (b), FRI_mid (c), and ΔFRI (d) |
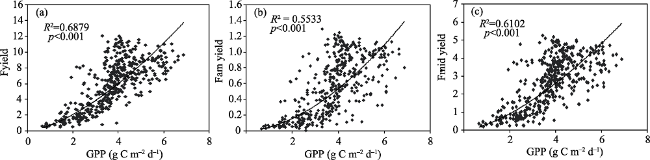
Fig. 6 Relationships of daily GPP (gC m-2 d-1) with Fyield (a), Fam-yield (b) and Fmid-yield (c). Fyield were calculated by dividing FRI by PAR from 8:30 a.m. to 17:00 p.m. Fam-yield were calculated by dividing FRI_am by PAR from 8:30 a.m. to 9:30 p.m. Fmid-yield were calculated by dividing FRI_mid by PAR from 11:30 a.m. to 13:30 p.m. |
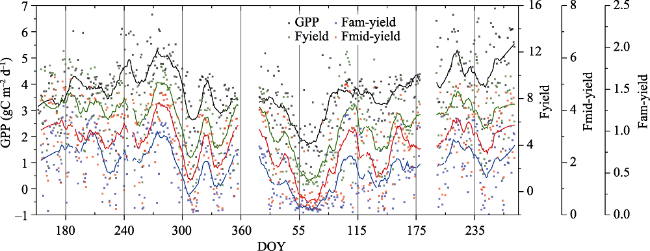
Fig. 7 Seasonal variations of GPP (gC m-2 d-1, black dots and lines), Fyield (red dots and lines), Fam-yield (blue dots and lines) and FRI_mid (green dots and lines) from 2014 (DOY 152, 1 June) to 2015 (DOY 273, 30 September). The solid lines indicate moving averages of 30 days. |
Table 1 Coefficients of correlation (R2) between GPP and various FRI and Fyield during different seasons |
| Index | FRI | FRI_am | FRI_mid | Fyield | Fam-yield | Fmid-yield |
|---|---|---|---|---|---|---|
| All seasons | 0.2243 | 0.3248 | 0.1332 | 0.6879 | 0.5533 | 0.6102 |
| Recovery-growth season | 0.1573 | 0.3021 | 0.0936 | 0.5974 | 0.4773 | 0.5163 |
| Senescence season | 0.5373 | 0.4876 | 0.4434 | 0.7924 | 0.6522 | 0.7353 |
Note: All correlations were significant (p<0.001, Pearsons correlation test). |
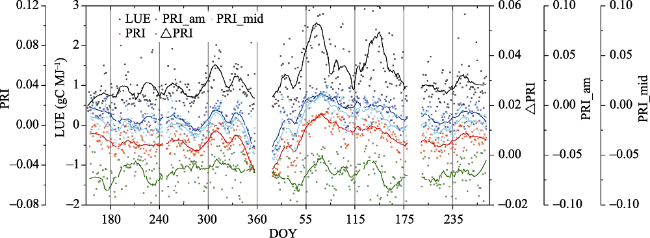
Fig. 8 Seasonal variation of LUE (gC MJ-1, black dots and lines), PRI (red dots and lines), PRI_am (blue dots and lines), PRI_mid (light blue dots and lines) and ΔPRI (green dots and lines) from 2014 (DOY 152, 1 June) to 2015 (DOY 273, 30 September). The solid lines indicate moving averages of 30 days. Daily average LUE and PRI were calculated using data observed from 8:30 a.m. to 17:00 p.m. each day. The average PRI_am were calculated using data observed from 8:30 a.m. to 9:30 a.m. The average PRI_mid were calculated using data observed from 11:30 a.m. to 13:30 p.m. ΔFRI were the differences between FRI_mid and FRI_am. ΔPRI were the differences between PRI_mid and PRI_am. |
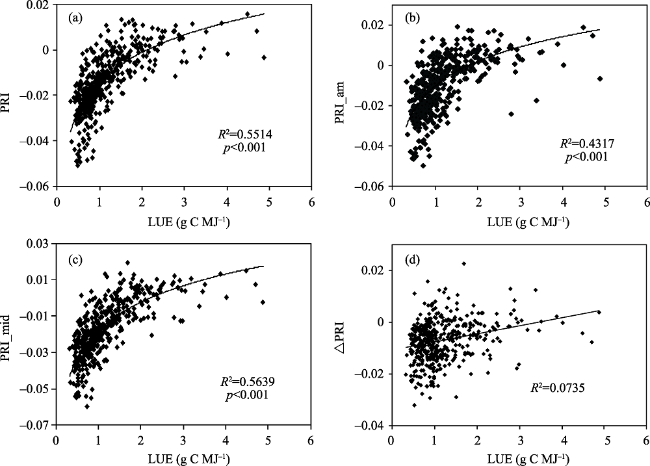
Fig. 9 Relationships of daily average LUE with PRI (a), PRI_am (b), PRI_mid (c) and ΔPRI (d) |
Table 2 Coefficients of correlation (R2) between LUE and PRI, PRI_am, and PRI_mid. during different seasons. |
| Index | PRI | PRI_am | PRI_mid |
|---|---|---|---|
| All seasons | 0.5514 | 0.4317 | 0.5639 |
| Recovery-growth season | 0.5370 | 0.3883 | 0.5181 |
| Senescence season | 0.8034 | 0.7001 | 0.7995 |
Note: All correlations were significant (p<0.001, Pearsons correlation test). |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}