
Journal of Resources and Ecology >
A Meta-analysis of the Effects of Warming and Elevated CO2 on Soil Microbes
Received date: 2018-06-05
Accepted date: 2018-09-12
Online published: 2019-01-28
Supported by
National Natural Science Foundation of China (31600432, 41571042)
The National Key Research Projects of China (2017YFA0604801)
The Youth Innovation Research Team Project of Key Laboratory of Ecosystem Network Observation and Modeling (LENOM2016Q0002)
Chinese Academy of Science Western Light Talents Program (Response of livestock carrying capability to climatic change and grazing in the alpine meadow of Northern Tibetan Plateau) and Tibet Science and Technology Major Projects of Pratacultural Industry.
Copyright
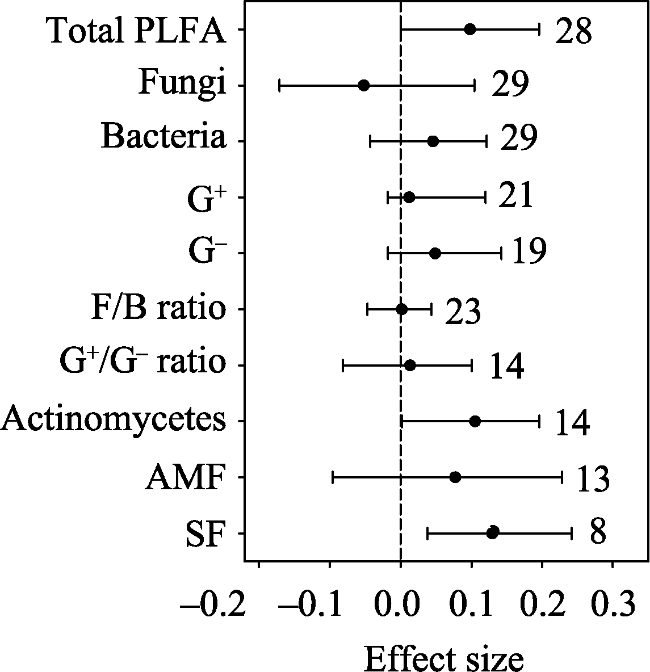
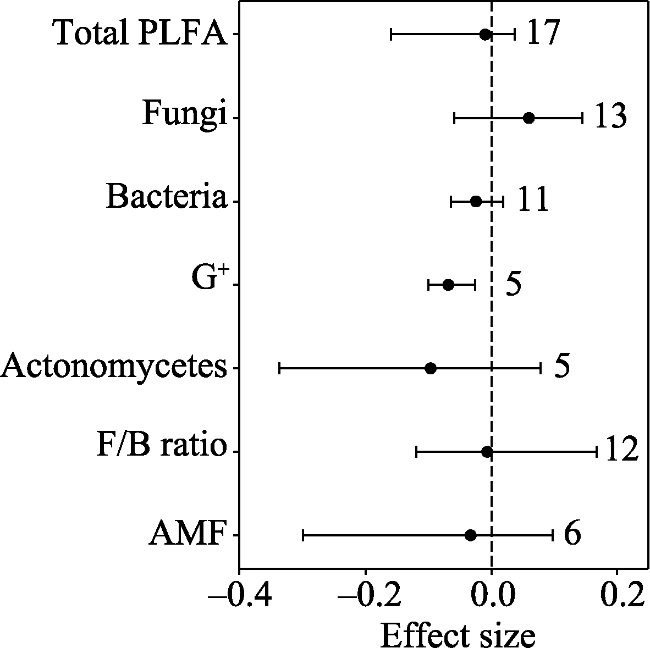
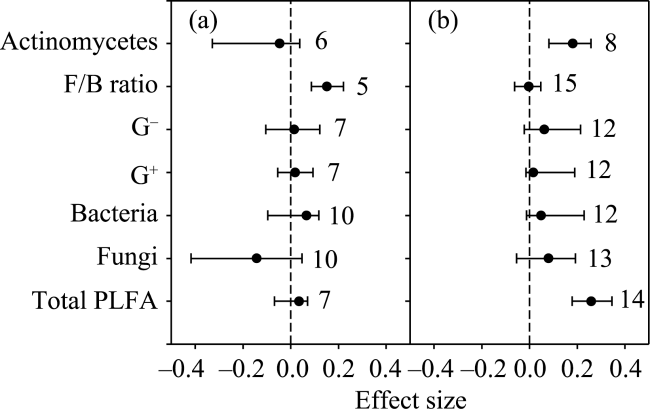
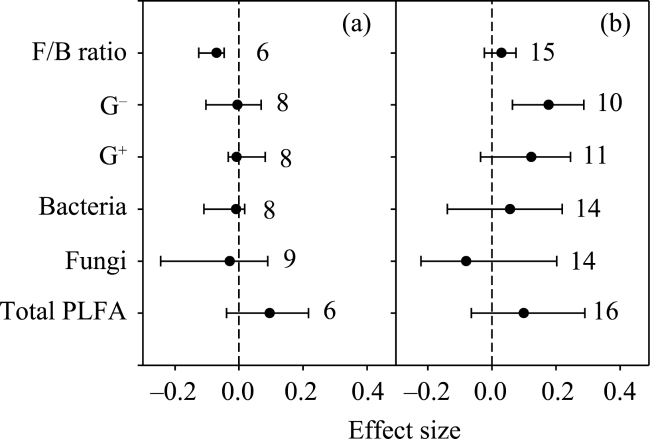
Soil microbes play important roles in terrestrial ecosystem carbon and nitrogen cycling. Climatic warming and elevated CO2 are two aspects of climatic change. In this study, we used a meta-analysis approach to synthesise observations related to the effects of warming and elevated CO2 on soil microbial biomass and community structure. Ecosystem types were mainly grouped into forests and grasslands. Warming methods included open top chambers and infrared radiators. Experimental settings included all-day warming, daytime warming and nighttime warming. Warming increased soil actinomycetes and saprotrophic fungi, while elevated CO2 decreased soil gram-positive bacteria (G+). Mean annual temperature and mean annual precipitation were negatively correlated with warming effects on gram-negative bacteria (G-) and total phospholipid fatty acid (PLFA), respectively. Elevation was positively correlated with the warming effect on total PLFA, bacteria, G+ and G-. Grassland exhibited a positive response of total PLFA and actinomycetes to warming, while forest exhibited a positive response in the ratio of soil fungi to bacteria (F/B ratio) to warming. The open top chamber method increased G-, while the infrared radiator method decreased the F/B ratio. Daytime warming rather than all-day warming increased G-. Our findings indicated that the effects of warming on soil microbes differed with ecosystem types, warming methods, warming times, elevation and local climate conditions.
FU Gang , ZHANG Haorui , LI Shaowei , SUN Wei . A Meta-analysis of the Effects of Warming and Elevated CO2 on Soil Microbes[J]. Journal of Resources and Ecology, 2019 , 10(1) : 69 -76 . DOI: 10.5814/j.issn.1674-764X.2019.01.009
Table 1 Basic information for the 28 studies included in the meta-analysis |
| Authors | Year | Journal | Title | MAT (℃) | MAP (mm) | Elevation (m) | Latitude | Longitude | Vegetation types |
|---|---|---|---|---|---|---|---|---|---|
| Wang et al. | 2011 | Chinese Journal of Applied and Environmental Biology | Microbial communities of alpine meadow soil in the Eastern Qinghai-Tibetan Plateau subjected to experimental warming and grazing | 1.1 | 752.4 | 3561 | 32.45 | 102.37 | Grassland |
| Schindlbacher et al. | 2011 | Soil Biology & Biochemistry | Experimental warming effects on the microbial community of a temperate mountain forest soil | 5.7 | 1480 | 910 | 47.58 | 11.64 | Forest |
| Andresen et al. | 2014 | PLoS ONE | Bacteria and fungi respond differently to multifactorial climate change in a temperate heathland, traced with 13C-glycine and FACE CO2 | 8 | 600 | 55.88 | 11.97 | Tundra | |
| Rinnan et al. | 2007 | Global Change Biology | Fifteen years of climate change manipulations alter soil microbial communities in a subarctic heath ecosystem | -0.4 | 245.5 | 450 | 68.35 | 18.82 | Tundra |
| Rousk et al. | 2013 | Global Change Biology | Investigating the long-term legacy of drought and warming on the soil microbial community across five European shrubland ecosystems | 56.38 | 10.95 | Shrubland | |||
| 46.88 | 19.72 | Shrubland | |||||||
| 40.60 | 8.15 | Shrubland | |||||||
| 52.40 | 5.92 | Shrubland | |||||||
| 53.05 | -3.47 | Shrubland | |||||||
| Zhou et al. | 2012 | Nature Climate Change | Microbial mediation of carbon-cycle feedbacks to climate warming | 16.3 | 914 | 34.98 | -97.52 | Grassland | |
| Zhao et al. | 2014 | Plant and Soil | Effects of experimental warming and nitrogen fertilization on soil microbial communities and processes of two subalpine coniferous species in Eastern Tibetan Plateau, China | 8.9 | 919.5 | 1820 | 31.68 | 103.88 | Forest |
| Zhang et al. | 2014 | PLoS ONE | Responses of soil microbial communities to experimental warming in alpine grasslands on the Qinghai-Tibet Plateau | -3.8 | 290.9 | 4635 | 34.82 | 92.93 | Grassland |
| Zhang et al. | 2011 | Soil Biology & Biochemistry | Soil microbial community changes and their linkages with ecosystem carbon exchange under asymmetrically diurnal warming | 2.1 | 383 | 1324 | 42.03 | 116.28 | Grassland |
| Shen et al. | 2014 | Pedosphere | Soil microbial responses to experimental warming and nitrogen addition in a temperate steppe of Northern China | 2.1 | 383 | 1324 | 42.03 | 116.28 | Grassland |
| Zhang et al. | 2013 | Oecologia | Soil microbial responses to warming and increased precipitation and their implications for ecosystem C cycling | 2.1 | 383 | 1324 | 42.03 | 116.28 | Grassland |
| Wang et al. | 2014 | Acta Ecologica Sinica | Effects of warming on soil microbial community structure in Changbai Mountain tundra | -7.3 | 1600 | 2028 | Tundra | ||
| Gutknecht et al. | 2012 | Global Change Biology | Microbial communities and their responses to simulated global change fluctuate greatly over multiple years | 37.67 | -122.37 | Grassland | |||
| Xu et al. | 2015 | Soil Biology & Biochemistry | Labile, recalcitrant, microbial carbon and nitrogen and the microbial community composition at two Abies faxoniana forest elevations under elevated temperatures | 2.7 | 813 | 3000 | Forest | ||
| Maestre et al. | 2015 | Frontiers in Microbiology | Warming reduces the cover and diversity of biocrust-forming mosses and lichens, and increases the physiological stress of soil microbial communities in a semi-arid Pinus halepensis plantation | 14.6 | 315 | 38.54 | -49.00 | Forest | |
| Zhang et al. | 2015 | European Journal of Soil Science | Depth-related responses of soil microbial communities to experimental warming in an alpine meadow on the Qinghai-Tibet Plateau | -3.8 | 383 | 4635 | 34.85 | 92.93 | Grassland |
| De Long et al. | 2016 | Ecosystems | Contrasting responses of soil microbial and nematode communities to warming and plant functional group removal across a post-fire boreal forest successional gradient | Forest | |||||
| Yoshitake et al. | 2015 | Ecological Research | Soil microbial response to experimental warming in cool temperate semi-natural grassland in Japan | 7.1 | 2128 | 36.13 | 137.42 | Grassland | |
| Authors | Year | Journal | Title | MAT (℃) | MAP (mm) | Elevation (m) | Latitude | Longitude | Vegetation types |
| Kao-Kniffin et al. | 2013 | Microbial Ecology | A microbial link between elevated CO2 and methane emissions that is plant species-specific | Grassland | |||||
| Feng et al. | 2010 | Global Change Biology | Altered microbial community structure and organic matter composition under elevated CO2 and N fertilization in the duke forest | 35.97 | -79.08 | Forest | |||
| Kasurinen et al. | 2005 | Global Change Biology | Below-ground responses of silver birch trees exposed to elevated CO2 and O3 levels during three growing seasons | 307.33 | 120 | 62.65 | 27.05 | Forest | |
| Kanerva et al. | 2008 | Soil Biology & Biochemistry | Changes in soil microbial community structure under elevated changes in soil microbial community structure under elevated tropospheric O3 and CO2 | 60.82 | 23.47 | Grassland | |||
| Ebersberger et al. | 2004 | Plant and Soil | Effects of long term CO2 enrichment on microbial community structure in calcareous grassland | 8.75 | 900 | 520 | 47.55 | 7.57 | Grassland |
| Janus et al. | 2005 | Microbial Ecology | Elevated atmospheric CO2 alters soil microbial communities associated with trembling aspen (Populus tremuloides) roots | 45.57 | -84.67 | Forest | |||
| Hagedorn et al. | 2013 | Soil Biology & Biochemistry | Nine years of CO2 enrichment at the alpine treeline stimulates soil respiration but does not alter soil microbial communities | 47.47 | 7.50 | Forest | |||
| Manninen et al. | 2010 | Soil Biology & Biochemistry | Plant and soil microbial biomasses in Agrostis capillaris and Lathyrus pratensis monocultures exposed to elevated O3 and CO2 for three growing seasons | 60.82 | 23.47 | Grassland | |||
| Chung et al. | 2007 | Global Change Biology | Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function | Grassland | |||||
| Guenet et al. | 2012 | Geoderma | The impact of long-term CO2 enrichment and moisture levels on soil microbial community structure and enzyme activities | Grassland |
MAT mean annual temperature; MAP mean annual precipitation. |
Fig. 1 Warming effects on soil microbial biomass |
Fig. 2 Elevated CO2 effects on soil microbial biomass |
Table 2 Relationships between warming effects on soil microbial biomass and relevant variables |
| Variables | Slope | p | QM | QE | QT | n |
|---|---|---|---|---|---|---|
| Warming duration | ||||||
| Total PLFA | -0.01 | 0.633 | 0.23 | 29.21 | 29.44 | 28 |
| Fungi | 0.01 | 0.568 | 0.33 | 24.81 | 25.13 | 28 |
| Bacteria | 0.01 | 0.394 | 0.73 | 29.67 | 30.39 | 29 |
| G+ | 0.03 | 0.417 | 0.66 | 20.04 | 20.69 | 20 |
| G- | 0.01 | 0.849 | 0.04 | 18.21 | 18.25 | 18 |
| F/B ratio | -0.01 | 0.471 | 0.52 | 18.79 | 19.31 | 22 |
| Warming magnitude | ||||||
| Total PLFA | -0.02 | 0.819 | 0.05 | 27.44 | 27.49 | 24 |
| Fungi | 0.05 | 0.517 | 0.42 | 22.01 | 22.43 | 26 |
| Bacteria | 0.01 | 0.887 | 0.02 | 27.36 | 27.38 | 26 |
| G+ | 0.01 | 0.904 | 0.01 | 22.38 | 22.39 | 21 |
| G- | 0.03 | 0.580 | 0.31 | 19.21 | 19.52 | 19 |
| F/B ratio | -0.01 | 0.815 | 0.05 | 16.16 | 16.21 | 19 |
| Mean annual air temperature | ||||||
| Total PLFA | -0.01 | 0.451 | 0.57 | 18.44 | 19.00 | 20 |
| Fungi | -0.01 | 0.366 | 0.82 | 14.80 | 15.62 | 20 |
| Bacteria | -0.01 | 0.269 | 1.22 | 17.68 | 18.90 | 21 |
| G+ | -0.01 | 0.166 | 1.92 | 22.23 | 24.16 | 20 |
| G- | -0.02 | 0.014 | 6.09 | 19.75 | 25.84 | 18 |
| F/B ratio | -0.0021 | 0.633 | 0.23 | 16.55 | 16.78 | 19 |
| Mean annual precipitation | ||||||
| Total PLFA | -0.0002 | 0.019 | 5.54 | 24.16 | 29.70 | 19 |
| Fungi | -0.0002 | 0.077 | 5.15 | 19.67 | 24.82 | 19 |
| Bacteria | -0.0001 | 0.244 | 1.36 | 19.22 | 20.58 | 20 |
| G+ | -0.0002 | 0.109 | 2.57 | 19.34 | 21.91 | 20 |
| G- | -0.0002 | 0.208 | 1.58 | 18.43 | 20.01 | 18 |
| F/B ratio | 0.0000 | 0.597 | 0.28 | 14.33 | 14.61 | 18 |
| Elevation | ||||||
| Total PLFA | 0.0001 | 0.031 | 4.63 | 16.81 | 21.45 | 17 |
| Fungi | 0.00 | 0.366 | 0.82 | 15.94 | 16.76 | 17 |
| Bacteria | 0.0001 | 0.018 | 5.63 | 15.45 | 21.09 | 17 |
| G+ | 0.0001 | 0.010 | 6.63 | 19.30 | 25.93 | 17 |
| G- | 0.0001 | 0.034 | 4.51 | 19.65 | 24.16 | 16 |
Note: Slope: regression coefficients; P: the statistical probability; n: the number of the observations used in the meta-analysis; QT: Total heterogeneity of the effect sizes of treatments among all studies; QM: the variation that can be explained by the continuous randomized-effects model; QE: The residual error; A significant slope (P < 0.05) indicates that an independent variable can explain the variation among effect sizes. G+: gram-positive bacteria, G-: gram-negative bacteria, F/B ratio: the ratio of soil fungi to bacteria. |
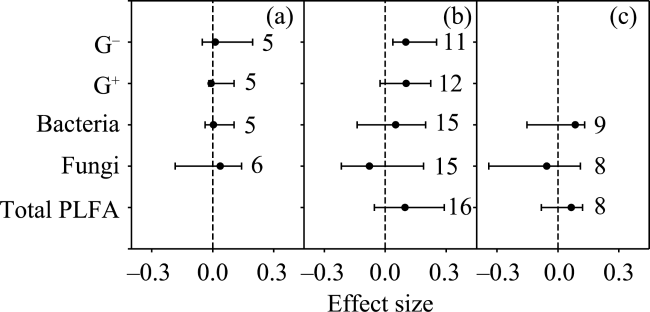
Fig. 3 Warming effects on soil microbial biomass for (a) forest and (b) grassland |
Fig. 4 Effects of (a) infrared radiator and (b) open top chamber on soil microbial biomass |
Fig. 5 Effects of (a) all-day, (b) daytime and (c) nighttime warming on soil microbial biomass |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}